
Превращение веществ и энергии в процессе диссимиляции проходит в три этапа.
1 этап. Подготовительный: сложные органические вещества под действием пищеварительных ферментов распадаются на простые, при этом выделяется только тепловая энергия. Белки расщепляются до аминокислот, жиры до глицерина и жирных кислот, крахмал до глюкозы.
2 этап. Гликолиз (бескислородный): происходит в цитоплазме, ферменты в нем не участвуют. Глюкоза расщепляется до молочной кислоты, при этом 40% выделившейся энергии идет на синтез АТФ.
3 этап. Гидролиз (Кислородный): осуществляется в митохондриях. Процесс происходит в матриксе и на внутренних мембранах. Процесс ферментативный. Расщеплению подвергается молочная кислота. Диоксид углерода выделяется из митохондрии в окружающую среду. Атом водорода (Н) включается в цепь ферментативных реакций в результате которых синтезируется АТФ.
Реакции протекают в следующей последовательности:
1. Атом водорода с помощью ферментов переносчиков поступает во внутреннюю мембрану митохондрий, образующую кристы, где и окисляется.
2. Протон водорода выносится переносчиками на наружную поверхность мембраны крист. Для протонов эта мембрана непроницаема, поэтому она накапливаются в межмембранном пространстве, образуя протонный резервуар.
3. Электроны водорода переносятся на внутреннюю поверхность мембраны крист и тут же присоединяются к кислороду с помощью фермента оксидазы, образуя отрицательно заряженный активный кислород.
4. Разноименно заряженные ионы по обе стороны мембраны создают электрическое поле, и когда разность потенциалов достигает 200 мВ, начитает действовать протонный канал. Он возникает в молекулах ферментов АТФ-синтетазы, которые встроены во внутреннюю мембрану, образующую кристы.
5. Через протонный канал протоны водорода устремляются внутрь митохондрии, воздавая высокий уровень энергии, большая часть который идет на синтез АТФ, а протоны,
взаимодействуя с активным кислородом, образуют воду и молекулярный кислород. Таким образом, кислород, поступающий в митохондрии в процессе дыхания, необходим для присоединения протонов водорода. При его отсутствии весь процесс в митохондриях прекращается, так как перестает функционировать электронно-транспортная цепь. Общая формула реакций третьего этапа:
Всего в процессе расщепления молекулы глюкозы образуется 38 молекул АТФ.
Статья на конкурс «био/мол/текст»: «Только ленивый не занимался митохондриями!» — сказал один профессор. И действительно, митохондрии — очень популярный объект исследования, ведь в них происходит множество сложнейших биохимических и биофизических процессов, обусловливающих широкий набор функций данных органелл. Но, несмотря на активность исследователей, многие механизмы этих процессов и свойства отдельных компонентов митохондрий остаются загадкой. Это связано в первую очередь с отсутствием подходящих неинвазивных методик. В ходе междисциплинарного проекта, проводимого группой биофизики клетки Биологического ф-та МГУ и лабораторией неорганического материаловедения химического ф-та МГУ совместно с коллегами из Германии и Дании, удалось создать методику на основе спектроскопии гигантского комбинационного рассеяния для селективного исследования цитохрома с непосредственно в живых митохондриях. Статья опубликована в журнале Scientific reports.
О, эти спектры! О, эти пики!В тайны молекул можем мы заглянуть.Так и работаем мы в этом ЛИКе*,И не напрасно избрали свой путь!
* — Лаборатория искусственного климата (ЛИК)— прежнее название корпуса каф. Биофизики биофака МГУ.
Жалюзи опущены. Выключен свет. Мы погружены во мрак. Капля мутной жидкости падает на серебряную подложку. Вспышка зелёного света. 30 секунд. Спектр. Так начинается наш длинный день. А теперь обо всём по порядку.
- Как всё устроено
- Что такое конформация гема и почему она важна?
- Как увидеть конформацию гема и оценить Еred/ox?
- К митохондриям от чистого сердца
- Усиливая сигнал
- Междисциплинарность — залог успеха
- Долгожданный спектр ГКР митохондрий
- Итоги
- Общая морфология.
- Ультраструктура митохондрий.
- Функции митохондрий.
- Окислительное фосфорилирование у бактерий.
- Увеличение числа митохондрий.
- Авторепродукция митохондрий.
- Хондриом.
Как всё устроено
* — Окислительно-восстановительный потенциал — синоним редокс-потенциала (от англ. reduction — восстановление, oxidation — окисление).
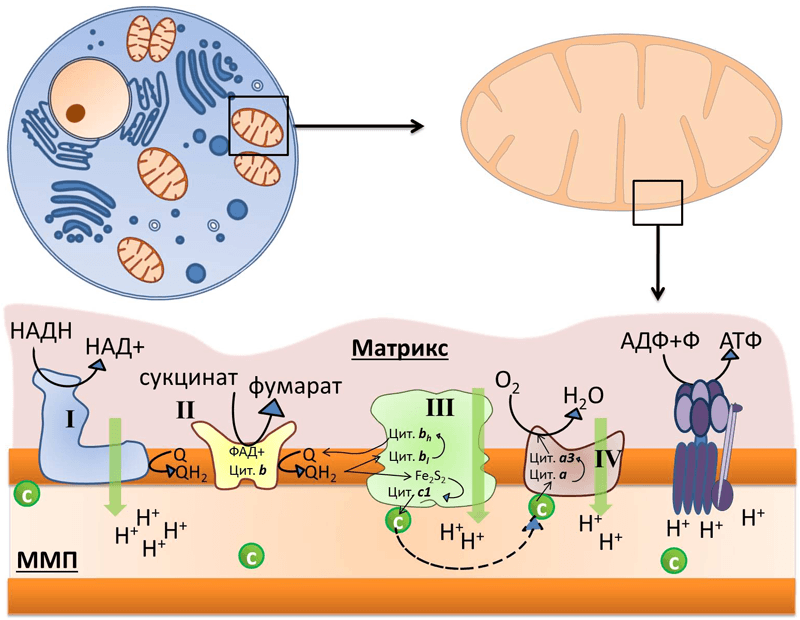
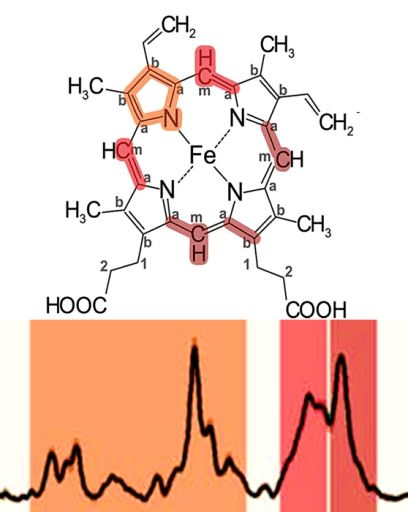
Рисунок 1. Во внутренней мембране митохондрий локализованы комплексы дыхательной цепи: I — НАДН-дегидрогеназа, II — сукцинат-дегидрогеназа, III — цитохром b–c1-комплекс, или цитохром с-редуктаза, IV — цитохром а–а3, или цитохромоксидаза, и фермент АТФ-синтаза. Q — убихинон, «с» в зелёном кружке — цитохром с. Чёрными стрелками показан транспорт электрона, пунктирной стрелкой — диффузия цитохрома с от комплекса III к IV, зелеными стрелками — перенос протона.
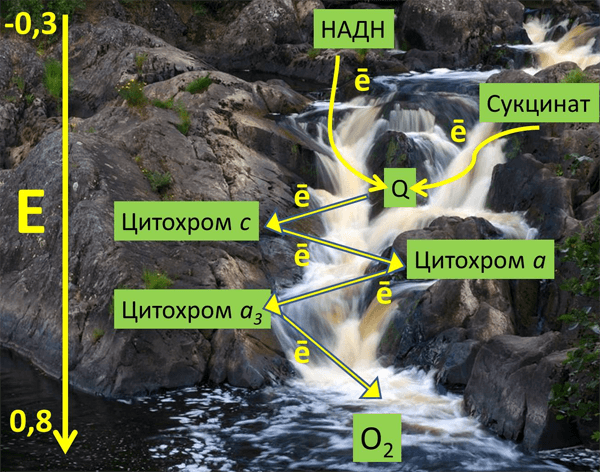
Рисунок 2. Метафорическое изображение среднеточечных редокс-потенциалов различных переносчиков ЭТЦ митохондрий в энергетической шкале (вольт) на фоне реки Тохмайоки в Карелии.
Что такое конформация гема и почему она важна?
Если приглядеться к комплексам дыхательной цепи (рис. 1), то в трёх из них можно увидеть цитохромы — гем-содержащие белки, которые являются кофакторами, участвующими в переносе электронов. А ещё один цитохром диффундирует в ММП. Всего в митохондриях имеется три типа цитохромов: а, b и с. Они очень похожи и отличаются только боковыми радикалами (рис. 3).
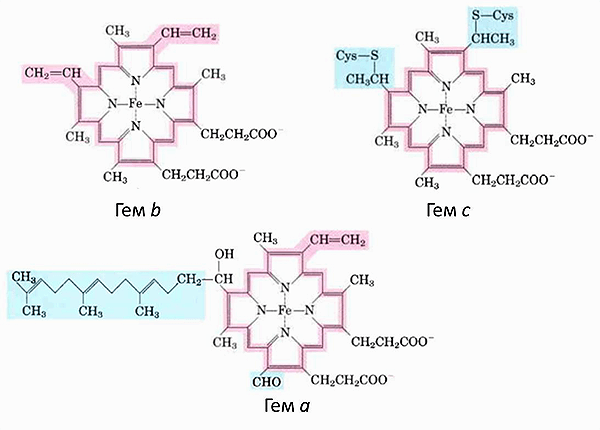
Возникает вопрос, как понять, в какой конформации и в каком редокс-состоянии находится гем цитохрома с в митохондриях?
Как увидеть конформацию гема и оценить Еred/ox?
Напрямую получение информации о конформации и редокс-состоянии цитохрома с в интактной (целой и функционирующей) митохондрии — это крайне сложная задача, ведь его конформация постоянно меняется при работе ЭТЦ, он диффундирует и взаимодействует с мембранными комплексами. Наличие метода, который бы позволял это сделать, значительно расширило бы наши знания о влиянии цитохрома с на активность дыхательной цепи и его вкладе в развитие митохондриальных патологий.
К митохондриям от чистого сердца
Однако у цитохромов есть одна особенность — спектр КР интенсивен только для восстановленных их форм, тогда как спектр окисленных состояний обладает очень низкой интенсивностью. Поэтому основным критерием определения восстановленности цитохромов является интенсивность сигнала. Но если мы ориентируемся только на интенсивность, то можем потерять важную информацию об изменениях в исследуемой системе! Ведь интенсивность зависит и от количества молекул, и от выраженности тех или иных колебаний, и от других параметров. К тому же, для определения точного положения пиков большинство исследователей, имеющих дело с цитохромами, вынуждены искусственно восстанавливать образцы, что заведомо нарушает их нативность.
Усиливая сигнал
Идея пришла от О.В. Сосновцевой, профессора Копенгагенского университета, с которой наша лаборатория сотрудничает не первый год. Она даже предложила «подсобить» с митохондриями, так как с ними работали в соседней лаборатории, и они готовы были поделиться.
Междисциплинарность — залог успеха
В 2010 году, когда интерес к нанотехнологиям был на высоте, академик Юрий Дмитриевич Третьяков, в то время декан факультета наук о материалах (ФНМ) МГУ, решил начать межфакультетское сотрудничество с биологами для проведения совместных исследований методом ГКР. В итоге сотрудничество было установлено с лабораторией биофизики клетки под руководством Г.В. Максимова. Так наша лаборатория познакомилась с заместителем декана ФНМ химиком-материаловедом член-корр. Е.А. Гудилиным. Совместно с группой Гудилина начались активные исследования по применению ГКР в биологии.
Дело в том, что ГКР — метод непростой и требует исключительно междисциплинарного подхода. Каждый биообъект уникален, и чтобы регистрировать от него сигнал ГКР, нужно разрабатывать уникальные наноструктуры, которые бы усиливали сигнал от нужного объекта, не повреждали его и выдерживали «атаку» физиологических многокомпонентных буферов.
Раз это удалось сделать для эритроцитов, то можно сделать и для митохондрий!
Долгожданный спектр ГКР митохондрий
Жалюзи опущены. Выключен свет. Комната погружена во мрак. Капля суспензии падает на серебряную подложку. Вспышка зелёного света. 30 секунд. Спектр. Тот самый долгожданный спектр ГКР от изолированных сердечных митохондрий был получен! Оставалось только понять, от каких именно структур в митохондриях исходит сигнал.
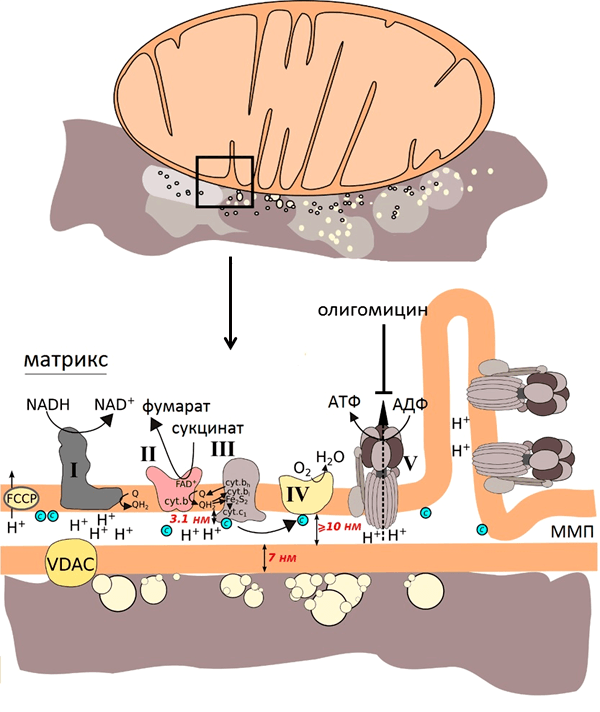
С учётом размеров компонентов митохондрий (рис. 7) и того, что усиление наблюдается на расстоянии нескольких нанометров от наноструктур, можно было предположить, что спектр ГКР митохондрий будет преимущественно спектром цитохрома с, так как этот цитохром наиболее близко подходит ко внешней мембране митохондрий и, соответственно, к наноструктурам. В то же время остальные цитохромы закреплены в комплексах внутренней мембраны.
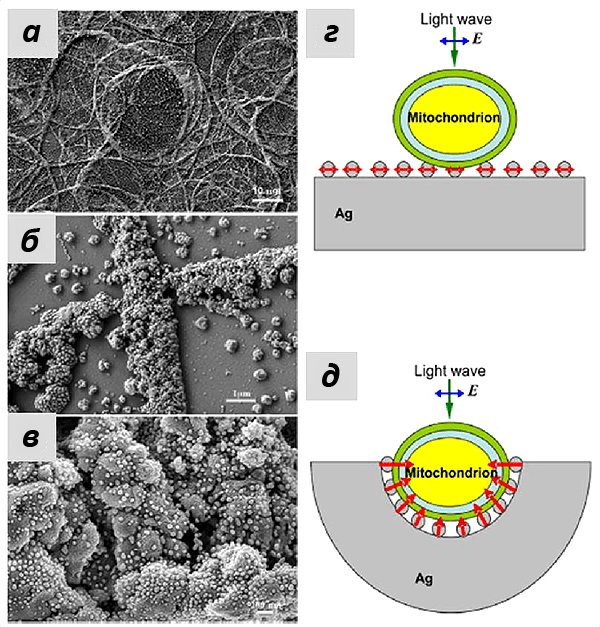
Это утверждение также подтверждалось при моделировании эффектов усиления КР серебряными наноструктурами, которые были использованы в работе. Группа наших коллег из лазерного центра Ганновера показала, что сложная морфология подложки со множеством углублений, в которые могут попадать митохондрии, позволило получать усиление на большом расстоянии (более 10 нм). А иерархическое устройство самих наноструктур увеличивало число мест контакта с мембраной митохондрий и, следовательно, число молекул цитохрома с, от которых можно было зарегистрировать сигнал ГКР. Если использовать те же наночастицы серебра, просто присоединенные к плоской подложке, то особого усиления не произойдет, что и подтверждалось в эксперименте (рис. 8).
И действительно, полученный спектр ГКР митохондрий соответствовал спектрам цитохрома с! При использовании зелёного лазера в качестве возбуждающего света можно регистрировать сигнал только от цитохромов b и с, но не от цитохрома а, что облегчает задачу расшифровки спектров. Несмотря на схожесть структуры цитохромов типа b и с, они имеют ключевые пики на спектре, благодаря которым их нельзя перепутать (рис. 9). Таким образом, используемые наноструктуры давали усиление на достаточно большом расстоянии, чтобы зарегистрировать спектры от цитохрома с, но недостаточно большом, для того чтобы увидеть пики цитохромов b. И это как раз то, что нужно! Благодаря методу ГКР теперь можно исследовать селективно редокс-состояние и конформацию именно цитохрома с в живых функционирующих митохондриях.
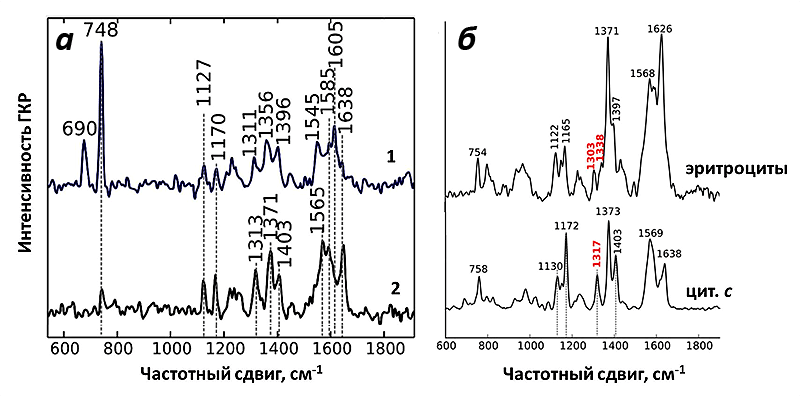
Как и ожидалось, спектры ГКР цитохрома с митохондрий оказались очень чувствительны к изменению его окислительно-восстановительного состояния. Для этого было исследовано два воздействия: внесение протонофора FCCP и ингибитора АТФ-синтазы олигомицина.
Таким образом, спектры ГКР митохондрий, являясь спектрами исключительно цитохрома с, оказались чувствительны к изменениям его конформации и редокс-состояния в процессе работы митохондрий.
Схема всей работы в одном ролике
Итоги
Спектроскопия гигантского комбинационного рассеяния позволяет многократно усилить сигнал КР от молекул вблизи наночастиц металла. Однако для проведения успешных экспериментов необходимо учитывать свойства как биологического объекта, так и наноструктур. «Ключевым моментом нашего достижения стал междисциплинарный подход к работе, в которую были вовлечены биологи, химики и физики», — поясняет одна из главных авторов проекта к.б.н. Надежда Браже, ставшая осенью 2015 года лауреатом премии L’Oreal Unesco для женщин в науке. — «Результатом такого подхода стало создание уникальной методики селективного определения редокс-состояния и конформации цитохрома с в живых функционирующих митохондриях, помещенных на специальную наноструктурированную поверхность. Разработанная методика поможет восполнить пробелы наших знаний о свойствах и поведении переносчиков электрона в митохондриях, а также может быть использована для разработки диагностических тестов для раннего выявления патологий митохондрий, чем и планируется заниматься в ближайшее время».
Общая морфология.
Митохондрии или хондриосомы (от греч. mitos – нить, chondrion – зернышко, soma – тельце) представляют собой гранулярные или нитевидные органеллы, присутствующие в цитоплазме простейших, растений и животных. Митохондрии можно наблюдать в живых клетках, так как они обладают достаточно высокой плотностью. В живых клетках митохондрии могут двигаться, перемещаться, сливаться друг с другом.
У разных видов размеры митохондрий очень непостоянны, так же как изменчива их форма (рис. 199). Все же у большинства клеток толщина этих структур относительно постоянна (около 0,5 мкм), а длина колеблется, достигая у нитчатых форм до 7-60 мкм.
Изучение величины и числа митохондрий не такое простое дело. Это связано с тем, что размеры и число митохондрий, которые видны на ультратонких срезах, не соответствуют реальности.
Обычные же подсчеты показывают, что на печеночную клетку приходится около 200 митохондрий. Это составляет более 20% от общего объема цитоплазмы и около 30-35% от общего количества белка в клетке. Площадь поверхности всех митохондрий печеночной клетки в 4-5 раз больше поверхности ее плазматической мембраны. Больше всего митохондрий в ооцитах (около 300000) и у гигантской амебы Chaos chaos (до 500000).
В клетках зеленых растений число митохондрий меньше, чем в клетках животных, так как часть их функций могут выполнять хлоропласты.
В спермиях часто присутствуют гигантские митохондрии, спирально закрученные вокруг осевой части жгутика. Отсутствуют митохондрии у кишечных энтамеб, живущих в условиях анаэробиоза, и у некоторых других паразитических простейших.
Локализация митохондрии в клетках различна. Обычно митохондрии скапливаются вблизи тех участков цитоплазмы, где возникает потребность в АТФ, образующейся в митохондриях. Так, в скелетных мышцах митохондрии находятся вблизи миофибрилл. В сперматозоидах митохондрии образуют спиральный футляр вокруг оси жгутика; вероятно, это связано с необходимостью использования АТФ для движения хвоста сперматозоида. Аналогичным образом у простейших и в других клетках, снабженных ресничками, митохондрии локализуются непосредственно под клеточной мембраной у основания ресничек, для работы которых необходим АТФ. В аксонах нервных клеток митохондрии располагаются около синапсов, где происходит процесс передачи нервного импульса. В секреторных клетках, которые синтезируют большие количества белков, митохондрии тесно связаны с зонами эргастоплазмы; вероятно, они поставляют АТФ для активации аминокислот и синтеза белка на рибосомах.
Ультраструктура митохондрий.
Митохондрии независимо от их величины или формы имеют универсальное строение, их ультраструктура однообразна. Митохондрии ограничены двумя мембранами (рис. 205). Наружняя митохондриальная мембрана отделяет ее от гиалоплазмы, она имеет ровные контуры, не образует впячиваний или складок, толщина составляет около 7 нм. На нее приходится около 7% от площади всех клеточных мембран. Мембрана не связана ни с какими другими мембранами цитоплазмы, замкнута сама на себя и представляет собой мембранный мешок. Наружнюю мембрану от внутренней отделяет межмембранное пространство шириной около 10-20 нм. Внутренняя мембрана (толщиной около 7 нм) ограничивает собственно внутреннее содержимое митохондрии, ее матрикс или митоплазму. Внутренняя мембрана митохондрий образовывает многочисленные впячивания внутрь митохондрий. Такие впячивания чаще всего имеют вид плоских гребней, или крист.
Общая поверхность внутренней мембраны митохондрии в печеночной клетке составляет примерно треть поверхности всех клеточных мембран. Митохондрии клеток сердечной мышцы содержат втрое больше крист, чем печеночные митохондрии что отражает различия в функциональных нагрузках митохондрий разных клеток. Расстояние между мембранами в кристе составляет около 10-20 нм.
Митохондриальные кристы, отходящие от внутренней мембраны и простирающиеся в сторону матрикса, не перегораживают полностью полость митохондрии и не нарушают непрерывности заполняющего ее матрикса.
Ориентация крист по отношению к длинной оси митохондрии различна для разных клеток. Ориентация может быть перпендикулярная (клетки печени, почек) крист; в сердечной мышце наблюдается продольное расположение крист. Кристы могут ветвиться или образовывать пальцевидные отростки, изгибаться и не иметь выраженной ориентации (рис. 208). У простейших, одноклеточных водорослей, в некоторых клетках высших растений и животных выросты внутренней мембраны имеют вид трубок (трубчатые кристы).
Матрикс митохондрий имеет тонкозернистое гомогенное строение, в котором выявляются молекулы ДНК в виде тонких собранных в клубок нитей (около 2-3 нм) и митохондриальные рибосомы имеющие форму гранул размером около 15-20 нм. Места отложения солей магния и кальция в матриксе образуют крупные (20-40 нм) плотные гранулы.
Функции митохондрий.
Митохондрии осуществляют синтез АТФ, происходящий в результате процессов окисления органических субстратов и фосфорилирования АДФ.
Начальные этапы окисления углеводов называются анаэробным окислением, или и происходят в гиалоплазме и не требуют участия кислорода. Субстратом окисления при анаэробном получении энергии служат гексозы и в первую очередь глюкоза; некоторые бактерии обладают свойством извлекать энергию, окисляяя пентозы, жирные кислоты или аминокислоты.
В глюкозе количество потенциальной энергии, заключенной в связях между атомами С, Н и О, составляет около 680 ккал на 1 моль (т.е. на 180 г глюкозы).
В живой клетке это огромное количество энергии освобождается в виде ступенчатого процесса, управляемого целым рядом окислительных ферментов, и не связано с переходом энергии химической связи в тепло, как при горении, а переходит в макроэнергетическую связь в молекулах АТФ, которые синтезируются при использовании освобождающейся энергии из АДФ и фосфата.
В процессе гликолиза происходит неполное окисление субстрата. В результате гликолиза глюкоза распадается до триоз, при этом тратятся 2 молекулы АТФ и синтезируются 4 молекулы АТФ. В конечном результате клетка получает всего 2 молекулы АТФ. В энергетическом отношении этот процесс малоэффективен, поэтому что из 680 ккал, заключающихся в связях 1 моля глюкозы, освобождается менее 10% энергии. Несмотря на это, анаэробное окисление, гликолиз, широко используется в живой природе. Он является основным поставляющим энергию процессом для многих микроорганизмов, некоторых кишечных паразитических анаэробных простейших, для клеток высших организмов на ранних стадиях эмбрионального развития, для многих опухолевых клеток, для клеток культуры ткани и др. Эритроциты млекопитающих, например, получают всю необходимую им энергию за счет гликолиза, так как у них нет митохондрий.
Образовавшиеся в результате гликолиза триозы, и в первую очередь пировиноградная кислота, в митохондриях вовлекаются в дальнейшее окисление. При этом происходит использование энергии расщепления всех химических связей, что приводит к выделению СО2, к потреблению кислорода и синтезу большого количества АТФ. Эти процессы связаны с окислительным циклом трикарбоновых кислот и с дыхательной цепью переноса электронов, где происходит фосфорилирование АДФ и синтез клеточного “топлива”, молекул АТФ (рис. 209).
В цикле трикарбоновых кислот (цикл Кребса, или цикл лимонной кислоты) образовавшийся в результате гликолиза пируват сначала теряет молекулу СО2 и, окисляясь до ацетата (двууглеродное соединение), соединяется с коферментом А. Затем ацетилкоэнзим А, соединяясь с оксалацетатом (четырехуглеродное соединение), образует шестиуглеродный цитрат (лимоную кислоту). Затем происходит цикл окисления этого шестиуглеродного соединения до четырехуглеродного оксалацетата, снова связывание с ацетилкоэнзимом А, и затем цикл повторяется. При этом окислении выделяются две молекулы СО2, а электроны, освободившиеся при окислении, переносятся на акцепторные молекулы коферментов (NAD-никотинамидадениндинуклеотид), которые вовлекают их далее в цепь переноса электронов. Следовательно, в цикле трикарбоновых кислот нет самого синтеза АТФ, а идет окисление молекул, перенос электронов на акцепторы и выделение СО2. Все описанные выше события внутри митохондрий происходят в их матриксе.
Окисление исходного субстрата приводит к выделению СО2 и воды, но при этом не выделяется тепловая энергия, как при горении, а образовываются молекулы АТФ. Они синтезируются другой группой белков, не связанных прямо с окислением. Во внутренних митохондриальных мембранах на поверхности мембран, смотрящих в матрикс, располагаются крупные белковые комплексы, ферменты, АТФ-синтетазы. В электронном микроскопе они видны в виде так называемых “грибовидных” телец сплошь выстилающие поверхность мембран, смотрящую в матрикс. Тельца имеют как бы ножку и головку, диаметром 8-9 нм. Следовательно, во внутренних мембранах митохондрий локализованы ферменты как окислительной цепи, так и ферменты синтеза АТФ (рис. 201б).
Выделяющаяся при транспорте электронов энергия запасается в виде градиента протонов на мембране. Оказалось, что при переносе электронов в митохондриальной мембране каждый комплекс дыхательной цепи направляет свободную энергию окисление на перемещение протонов (положительных зарядов) через мембрану, из матрикса в межмембранное пространство, что приводит к образованию разности потенциалов на мембране: положительные заряды преобладают в межмембранном пространстве, а отрицательные – со стороны матрикса митохондрий. При достижении разности потенциалов (220 мВ) белковый комплекс АТФ-синтетазы начинает транспортировать протоны обратно в матрикс, при этом превращает одну форму энергии в другую: образует АТФ из АДФ и неорганического фосфата. Так происходит сопряжение окислительных процессов с синтетическим, с фосфорилированием АДФ. Пока происходит окисление субстратов, пока происходит перекачка протонов через внутреннюю митохондриальную мембрану – идет сопряженный с этим синтез АТФ, т.е. происходит окислительное фосфорилирование.
Эти два процесса можно разобщить. При этом продолжается перенос электронов, как и окисление субстрата, но синтеза АТФ не происходит. В этом случае энергия, освобождающаяся при окислении переходит в тепловую энергию.
Окислительное фосфорилирование у бактерий.
У прокариотических клеток, способных к окислительному фосфорилированию, элементы цикла трикарбоновых кислот локализованы прямо в цитоплазме, а ферменты дыхательной цепи и фосфорилирования связаны с клеточной мембраной, с ее выпячиваниями, выступающими внутрь цитоплазмы, с так называемыми мезосомами (рис. 212). Надо отметить, что такие бактериальные мезосомы могут быть связаны не только с процессами аэробного дыхания, но и у некоторых видов участвовать в делении клеток, в процессе распределения ДНК по новым клеткам, в образовании клеточной стенки и т.д.
На плазматической мембране в мезосомах некоторых бактерий осуществляются сопряженные процессы как окисления так и синтеза АТФ. В электронном микроскопе во фракциях плазматических мембран бактерий обнаружены сферические частицы, аналогичные тем, которые были найдены в митохондриях эукариотических клеток. Таким образом, у бактериальных клеток, способных к окислительному фосфорилированию, плазматическая мембрана выполняет роль, аналогичную внутренней мембране митохондрий эукариотических клеток.
Увеличение числа митохондрий.
Митохондрии могут увеличивать свою численность особенно при делении клеток или при увеличении функциональной нагрузки клетки. Происходит постоянное обновление митохондрий. Например, в печени средняя продолжительность жизни митохондрий составляет около 10 дней.
Увеличение числа митохондрий происходит путем роста и деления предшествующих митохондрий. Это предположение было впервые высказано Альтманом (1893), описавшим митохондрии под термином “биобласты”. Удается наблюдать прижизненно деление, фрагментацию длинных митохондрий на более короткие путем перетяжки, что напоминает бинарный способ деления бактерий.
Реальное увеличение числа митохондрий путем деления установлено при изучении поведения митохондрий в живых клетках культуры ткани. В течение клеточного цикла митохондрии вырастают до нескольких мкм, а затем фрагментируются, делятся на более мелкие тельца.
Митохондрии могут сливаться друг с другом и размножаться по принципу: митохондрия от митохондрии.
Авторепродукция митохондрий.
Двумембранные органеллы обладают полной системой авторепродукции. В митохондриях и пластидах существует ДНК, на которой синтезируются информационные, трансферные и рибосомные РНК и рибосомы, осуществляющие синтез митохондриальных и пластидных белков. Однако, эти системы, хотя и автономны, но ограничены по своим возможностям.
ДНК в митохондриях представляет собой циклические молекулы без гистонов и тем самым напоминают бактериальные хромосомы. Размер их составляет около 7 мкм, в одну циклическую молекулу митохондрий животных входит 16-19 тыс. нуклеотидных пар ДНК. У человека митохондриальная ДНК содержит 16,5 тыс. н.п., она полностью расшифрована. Найдено, что митохондральная ДНК различных объектов очень однородна, отличие их заключается лишь в величине интронов и нетранскрибируемых участков. Все митохондриальные ДНК представляют множественные копии, собранными в группы, кластеры. Так в одной митохондрии печени крысы может содержаться от 1 до 50 циклических молекул ДНК. Общее же количество митохондриальной ДНК на клетку составляет около одного процента. Синтез митохондриальных ДНК не связан с синтезом ДНК в ядре.
Так же как и у бактерий митохондральная ДНК собрана в отдельную зону – нуклеоид, его размер составляет около 0, 4 мкм в диаметре. В длинных митохондриях может быть от 1 до 10 нуклеоидов. При делении длинной митохондрии от нее отделяется участок, содержащий нуклеоид (сходство с бинарным делением бактерий). Количество ДНК в отдельных нуклеоидах митохондрий может колебаться в 10 раз в зависимости от типа клеток.
В некоторых культурах в клетках от 6 до 60% митохондрий не имеют нуклеоида, что может объясняться тем, что деление этих органелл скорее связано с фрагментацией, а не с распределением нуклеоидов.
Как уже говорилось, митохондрии могут как делиться, так и сливаться друг с другом. При слиянии митохондрий друг с другом может происходить обмен их внутренними компонентами.
Важно подчеркнуть, что рРНК и рибосомы митохондрий и цитоплазмы резко отличны. Если в цитоплазме обнаруживаются 80s рибосомы, то рибосомы митохондрий растительных клеток принадлежат к 70s рибосомам (состоят из 30s и 50s субъединиц, содержат 16s и 23s РНК, характерные для прокариотических клеток), а в митохондриях клеток животных обнаружены более мелкие рибосомы (около 50s).
Рибосомные РНК митохондрий синтезируются на митохондриальных ДНК. В митоплазме на рибосомах идет синтез белков. Он прекращается, в отличие от синтеза на цитоплазматических рибосомах, при действии антибиотика хлорамфеникола, подавляющего синтез белка у бактерий.
На митохондриальном геноме синтезируются 22 транспортные РНК. Триплетный код митохондриальной синтетической системы отличен от такового, используемого в гиалоплазме. Несмотря на наличие казалось бы всех компонентов, необходимых для синтеза белков, небольшие молекулы митохондриальной ДНК не могут кодировать все митохондриальные белки, только лишь их небольшую часть. Так ДНК размером 15 тыс.н.п. может кодировать белки с суммарным молекулярным весом около 6х105. В это же время суммарный молекулярный вес белков частицы полного дыхательного ансамбля митохондрии достигает величины около 2х106.
Если учесть, что кроме белков окислительного фосфорилирования в митохондрии входят ферменты цикла трикарбоновых кислот, ферменты синтеза ДНК и РНК, ферменты активации аминокислот и другие белки, то видно, что, для того чтобы кодировать эти многочисленные белки и рРНК и тРНК, количества генетической информации в короткой молекуле митохондриальной ДНК явно не хватает. Расшифровка нуклеотидной последовательности митохондриальной ДНК человека показала, что она кодирует всего лишь 2 рибосомные РНК, 22 трансферных РНК и всего 13 различных полипептидных цепей.
В настоящее время доказано, что большая часть белков митохондрий находится под генетическим контролем со стороны клеточного ядра и синтезируется вне митохондрий. Большинство митохондриальных белков синтезируется на рибосомах в цитозоле. Эти белки имеют специальные сигнальные последовательности, которые узнаются рецепторами на внешней мембране митохондрий. Эти белки могут встраиваться в них (см. аналогию с мембраной пероксисом), а затем перемещаться на внутреннюю мембрану. Этот перенос происходит в точках контакта наружной и внутренней мембран, где такой транспорт отмечен. Большинство липидов митохондрий так же синтезируются в цитоплазме.
Все это говорит о эндосимбиотическом происхождении митохондрий, о том, что митохондрии представляют собой организмы типа бактерий, находящиеся в симбиозе с эукариотический клеткой.
Хондриом.
Совокупность всех митохондрий в одной клетке называется хондриомом. Она может быть различной в зависимости от типа клеток. Во многих клетках хондриом состоит из разрозненных многочисленных митохондрий, равномерно расположенных по всей цитоплазме или локализуются группами в местах интенсивной траты АТФ. В обоих этих случаях митохондрии функционируют поодиночке, их кооперативная работа, возможно, координируется какими-то сигналами из цитоплазмы. Существует и совершенно иной тип хондриома, когда вместо мелких одиночных разрозненных митохондрий в клетке располагается одна гигантская разветвленная митохондрия.
Такие митохондрии встречаются у одноклеточных зеленых водорослей (например у Chlorella). Они образуют сложную митохондриальную сеть или митохондриальный ретикулум (Reticulum miyochondriale). Согласно хемоосмотической теории биологический смысл появления такой гигантской разветвленной митохондриальной структуры, объединенной в одно целое своими внешними и внутренними мембранами заключается в том, что в любой точке поверхности внутренней мембраны такой разветвленной митохондрии может идти синтез АТФ, который будет поступать в любую точку цитоплазмы, где в этом есть необходимость.
В случае гигантских разветвленных митохондрий в любой ее точке может на внутренней мембране накопиться потенциал, достаточный для того, чтобы начался синтез АТФ. С этих позиций митохондриальный ретикулум представляет собой как бы электрический проводник, кабель, соединяющий отдаленные точки такой системы. Митохондриальный ретикулум оказался очень полезным не только для мелких подвижных клеток, таких как хлорелла, но и для более крупных структурных единиц таких как, например, миофибриллы в скелетных мышцах.
Известно, что скелетные мышцы состоят из массы мышечных волокон, симпластов, содержащих множество ядер. Длина таких мышечных волокон достигает 40 мкм, при толщине 0,1 мкм – это гигантская структура, содержащая великое множество миофибрилл, все из которых сокращаются одновременно, синхронно. Для сокращения к каждой единице сокращения, к миофибрилле, доставляется большое количество АТФ, которых обеспечивают митохондрии на уровне z-дисков. На продольных ультратонких срезах скелетных мышц в электронном микроскопе видны многочисленные округлые мелкие сечения митохондрий, располагающихся в соседстве с саркомерами. Мышечные митохондрии представляют собой не мелкие шарики или палочки, а как бы паукообразные структуры, отростки которых ветвятся и простираются на большие расстояния, иногда через весь поперечник мышечного волокна.
При этом разветвления митохондрий окружают каждую миофибриллу в мышечном волокне, снабжая их АТФ, необходимого для мышечного сокращения. Следовательно, в плоскости z-диска митохондрии представляют типичный митохондриальный ретикулум. Такой пласт или этаж митохондриального ретикулума повторяется дважды на каждый саркомер, а все мышечное волокно имеет тысячи поперечно расположенных “поэтажных” пластов митохондриального ретикулума. Между “этажами” вдоль миофибрилл располагаются нитчатые митохондрии, соединяющие эти митохондриальные пласты. Таким образом создана трехмерная картина митохондриального ретикулума, проходящего через весь объем мышечного волокна.
Далее было установлено, что между ответвлениями митохондриального ретикулума и нитевидными продольными митохондриями существуют специальные межмитохондриальные соединения или контакты (ММК). Они образованы плотно прилегающими наружными митохондриальными мембранами контактирующих митохондрий, межмембранное пространство и мембраны в этой зоне имеют повышенную электронную плотность. Через эти специальные образования происходит функциональное объединение соседних митохондрий и митохондриальных ретикулумов в единую, кооперативную энергетическую систему. Все миофибриллы в мышечном волокне сокращаются синхронно по всей их длине, следовательно, и поступление АТФ на любом участке этой сложной машины тоже должно происходить синхронно, а это может происходить лишь в том случае, если огромное количество разветвленных митохондрий-проводников будет связано друг с другом с помощью контактов.
О том, что межмитоходриальные контакты (ММК) участвуют в энергетическом объединении митохондрий друг с другом удалось на кардиомиоцитах, клетках сердечных мышц.
Хондриом клеток сердечной мышцы не образует ветвящихся структур, а представлен множеством небольших вытянутых митохондрий, располагающихся без особого порядка между миофибриллами. Однако, все соседние митохондрии стыкуются друг с другом с помощью митохондриальных контактов такого же типа, как в скелетной мышце, только их число очень велико: в среднем на одну митохондрию приходится 2-3 ММК, которые связывают митохондрии в единую цепь, где каждым звеном такой цепи (Streptio mitochondriale) является отдельная митохондрия.
Оказалось, что межмитохондриальные контакты (ММК), как обязательная структура сердечных клеток обнаружены в кардиомиоцитах как желудочков, так и предсердий всех позвоночных животных: млекопитающих, птиц, пресмыкающихся, амфибий и костистых рыб. Более того ММК были обнаружены (но в меньшем числе) в клетках сердца некоторых насекомых и моллюсков.
Количество ММК в кардиомиоцитах изменяется в зависимости от функциональной нагрузки на сердце. Число ММК увеличивается при повышении физических нагрузок животных и, наоборот, при падении нагрузки на сердечную мышцу происходит резкое сокращение числа ММК.






