- Как улучшить свои митохондрии?
- Чего боится митохондрия?
- Междисциплинарность — залог успеха
- Как всё устроено
- Как митохондрии управляют судьбой клетки?
- Этиология и патогенез митохондриальных заболеваний
- Немембранные органеллы. Рибосомы.
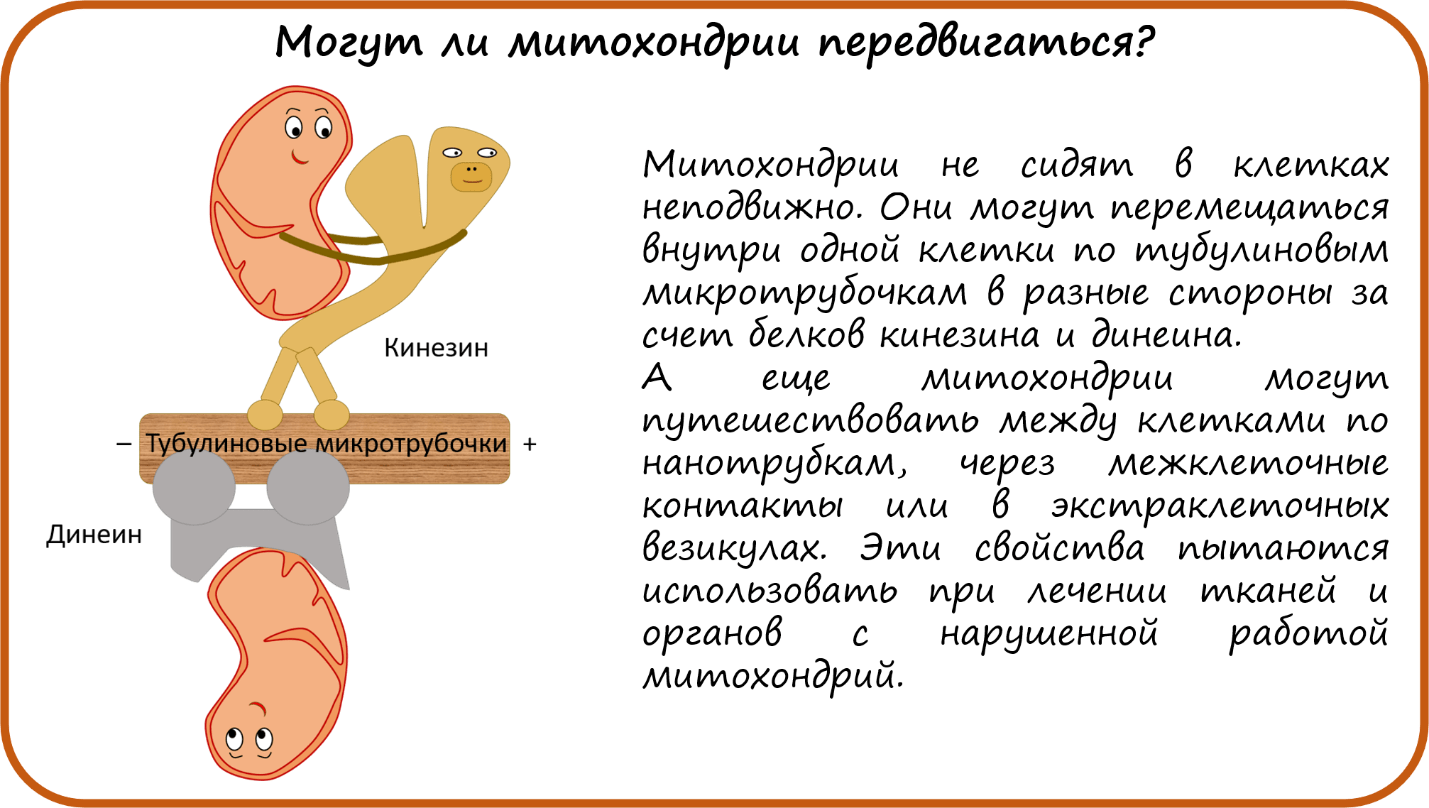
- Могут ли митохондрии передвигаться?
- Долгожданный спектр ГКР митохондрий
- К митохондриям от чистого сердца
- Усиливая сигнал
- На каком языке говорят митохондрии?
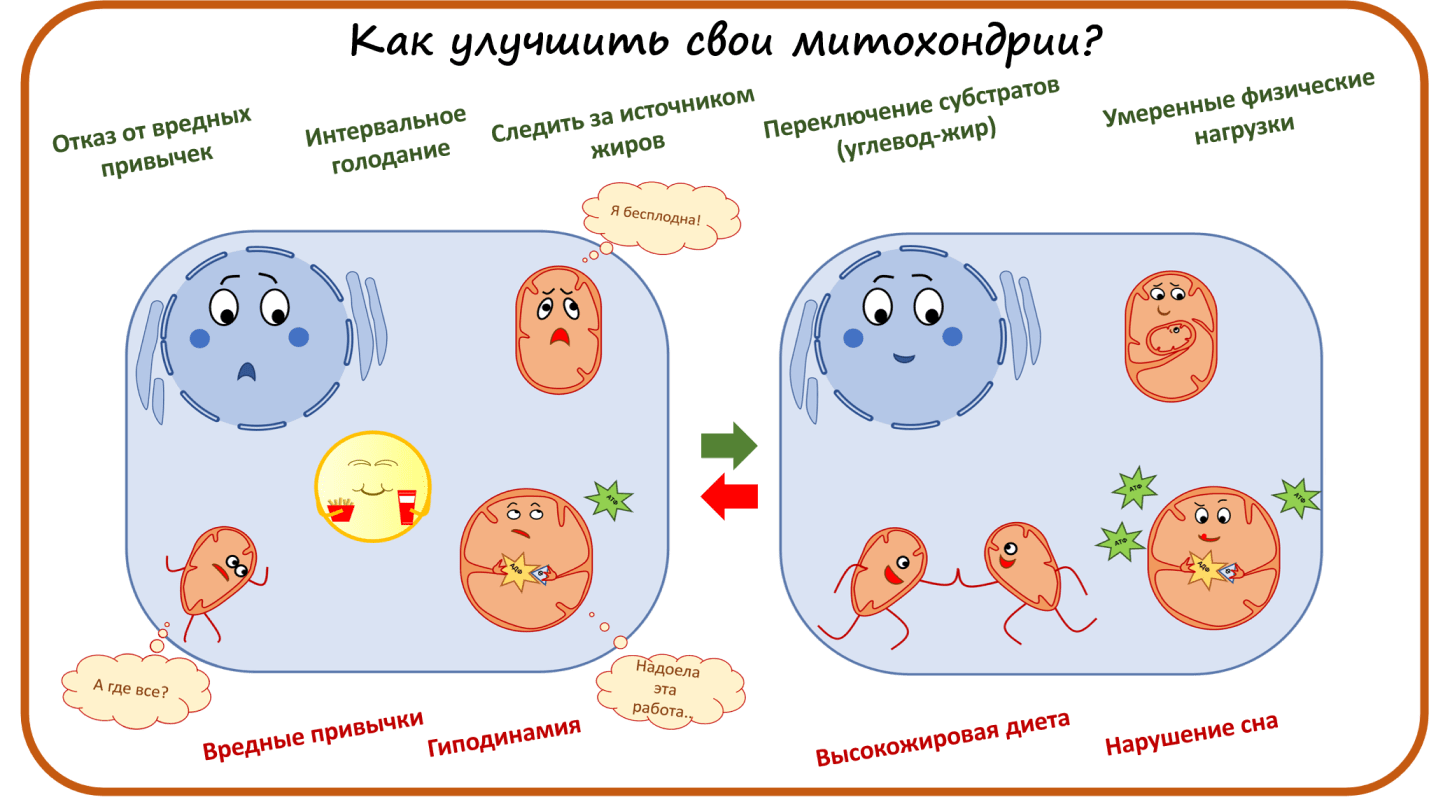
Как улучшить свои митохондрии?
В заключение хотелось бы сказать, что, хотя сейчас и разрабатывается множество перспективных препаратов, все же пока самый лучший способ держать митохондрии в тонусе — спорт, сбалансированное питание и отказ от вредных привычек!
Желаю, чтобы ваши митохондрии были здоровы и счастливы!

Видеолекцию автора можно посмотреть на YouTube.
Работа автора поддержана фондом РНФ (грант 21-74-00026)
Митохондрии
– это структуры палочковидной или
овальной формы (греч. mitos
–
нить,
chondros
– гранула). Они обнаружены во всех животных
клетках (исключая зрелые эритроциты):
у высших растений, у водорослей и
простейших. Отсутствуют они только у
прокариот бактерий.
Эти
органеллы впервые были обнаружены и
описаны в конце прошлого столетия
Альтманом. Несколько позже эти структуры
были названы митохондриями. В 1948 г.
Хогебум указал на значение митохондрий
как центра клеточного дыхания, а в 1949
г. Кеннеди и Ленинджер установили, что
в митохондриях протекает цикл
окислительного фосфорилирования. Так
было доказано, что митохондрии служат
местом генерирования энергии.
Митохондрии видны
в обычном световом микроскопе при
специальных методах окраски. В фазово
– контрастном микроскопе и в «темном
поле» их можно наблюдать в живых клетках.
Строение,
размеры, форма
митохондрий очень вариабельны. Это
зависит в первую очередь от функционального
состояния клеток. Например, установлено,
что в мотонейронах мух, летающих
непрерывно 2 часа, проявляется огромное
количество шаровидных митохондрий, а
у мух со склеенными крыльями число
митохондрий значительно меньше и они
имеют палочковидную форму (Л. Б. Левинсон).
По форме они могут быть нитевидными,
палочковидными, округлыми и гантелеобразными
даже в пределах одной клетки.
Митохондрии
локализованы в клетке, как правило, либо
в тех участках, где расходуется энергия,
либо около скоплений субстрата (например,
липидных капель), если таковые имеются.
Строгая ориентация
митохондрий обнаруживается вдоль
жгутиков сперматозоидов, в поперечно-полосатой
мышечной ткани, где они располагаются
вдоль миофибрилл, в эпителии почечных
канальцев локализуются во впячиваниях
базальной мембраны и т.д.
Количество
митохондрий в клетках имеет органные
особенности, например, в клетках печени
крыс содержится от 100 до 2500 митохондрий,
а в клетках собирательных канальцев
почки – 300, в сперматозоидах различных
видов животных от 20 до 72, у гигантской
амебы Chaos
chaos
их число достигает
500 000. Размеры митохондрий колеблются
от 1 до 10 мкм.
Ультрамикроскопическое
строение митохондрий однотипно,
независимо
от их формы и размера. Они покрыты двумя
липопротеидными мембранами: наружной
и внутренней. Между ними располагается
межмембранное пространство.
Впячивания
внутренней мембраны, которые вдаются
в тело митохондрий, называются кристами.
Расположение крист в митохондриях может
быть поперечным и продольным. По форме
кристы могут быть простыми и разветвленными.
Иногда они образует сложную сеть. В
некоторых клетках, например, в клетках
клубочковой зоны надпочечника кристы
имеют вид трубочек. Количество крист
прямо пропорционально интенсивности
окислительных процессов, протекающих
в митохондриях. Например, в митохондриях
кардиомиоцитов их в несколько раз
больше, чем в митохондриях гепацитов.
Пространство, ограниченное внутренней
мембраной, составляет внутреннюю камеру
митохондрий. В нем между кристами
находится митохондриальный матрикс —
относительно электронно плотное
вещество.
Белки внутренней
мембраны синтезируются миторибосомами,
а белки внешней мембраны – циторибосомами.
‘Наружная мембрана
митохондрий по многим показателям
сходна с мембранами ЭПС. Она бедна
окислительными ферментами. Немного их
и в мембранном пространстве. Зато
внутренняя мембрана и митохондриальный
матрикс буквально насыщены ими. Так, в
матриксе митохондрий сосредоточены
ферменты цикла Кребса и окисления жирных
кислот. Во внутренней мембране локализована
цепь переноса электронов, ферменты
фосфорилирования (образования АТФ из
АДФ), многочисленные транспортные
системы.
Кроме белка и
липидов, в состав мембран митохондрий
входит РНК, ДНК, последняя обладает
генетической специфичностью, и по своим
физико-химическим свойствам отличается
от ядерной ДНК.
При
электронно-микроскопических исследованиях
обнаружено, что поверхность наружной
мембраны покрыта мелкими шаровидными
элементарными частицами. Внутренняя
мембрана и кристы содержат подобные
элементарные частицы на «ножках», так
называемые грибовидные тельца. Они
-состоят из трех частей: головки
сферической формы (диаметр 90-100 А°), ножки
цилиндрической формы, длиной 5 нм и
шириной 3-4 нм, основания, имеющего размеры
4 на 11 нм. Головки грибовидных телец
связаны с фосфорилированием, затем
обнаружено, что головки содержат фермент,
обладающий АТФ-идной активностью.
В межмембранном
пространстве находится вещество,
обладающее более низкой электронной
плотностью, чем матрикс. Оно обеспечивает
сообщение между мембранами и поставляет
для ферментов, находящихся в обеих
мембранах, вспомогательные
катализаторы-коферменты.
В
настоящее время известно, что наружная
мембрана митохондрий хорошо проницаема
для веществ, имеющих низкий молекулярный
вес, в частности, белковых соединений.
Внутренняя мембрана митохондрий обладает
избирательной проницаемостью. Она
практически непроницаема для анионов
(Cl-1,
Br-1,
SO4-2,
HCO3-1,
катионов Sn+2,
Mg+2,
ряда cахаров
и большинства аминокислот, тогда как
Са2+,
Мп2+,
фосфат, многокарбоновые
кислоты
легко проникают через нее. Имеются
данные о наличии во внутренней мембране
нескольких переносчиков, специфических
к отдельным группам проникающих анионов
и катионов. Активный транспорт веществ
через мембраны осуществляется благодаря
использованию энергии АТФ-азной системы
или электрического потенциала,
генерируемого на мембране в результате
работы дыхательной цепи. Даже АТФ,
синтезированная в митохондриях, может
выйти с помощью переносчика (сопряженный
транспорт).
Матрикс
митохондрий представлен мелкозернистым
электронно-плотным веществом. В нем
располагаются миторибосомы, фибриллярные
структуры, состоящие из молекул ДНК и
гранул, имеющих диаметр более 200А◦
образованные
солями: Ca3(PO4),
Ba3(PO4)2,
Mg3(PO4).
Полагают, что гранулы служат резервуаром
ионов Са+2
и Мg+2.
Их количество увеличивается при изменении
проницаемости митохондриальных мембран.
Присутствие
в митохондриях ДНК обеспечивает участие
митохондрий в синтезе РНК и
специфических
белков, а также указывает на существование
цитоплазматической наследственности.
Каждая митохондрия содержит в зависимости
от размера одну или несколько молекул
ДНК (от 2 до 10). Молекулярный вес
митохондриальной ДНК около (30-40)*106
у простейших, дрожжей, грибов. У высших
животных около (9–10) *106.
Длина
ее у дрожжей примерно равна 5 мкм, у
растений – 30 мкм. Объем генетической
информации, заключенный в митохондриальной
ДНК, невелик: он состоит из 15-75 тыс. пар
оснований, которые могут кодировать в
среднем 25-125 белковых цепей с молекулярным
весом около 40000.
Митохондриальная
ДНК
отличается
от ядерной ДНК
рядом особенностей:
более высокой скоростью синтеза (в 5-7
раз), она более устойчива к действию
ДНК-азы, представляет собой двухкольцевую
молекулу, содержит больше гуанина и
цитозина, денатурируется при более
высокой температуре и легче
восстанавливается. Однако не все
митохондриальные белки синтезируются
митохондриальной
системой. Так, синтез цитохрома С и
других
ферментов
обеспечивается информацией, содержащейся
в ядре. В матриксе митохондрий локализованы,
витамины А, В2,
В12,
К, Е,
а
также гликоген.
Функция
митохондрий
заключается
в образовании энергии, необходимой для
жизнедеятельности клеток. Источником
энергии в клетке могут служить различные
соединения: белки, жиры, углеводы. Однако
единственным субстратом, который
немедленно включается в энергетические
процессы, является глюкоза.
Биологические
процессы, в результате которых в
митохондриях образуется энергия, можно
подразделить на 3 группы: I
группа – окислительные реакции, включающие
две фазы: анаэробную (гликолиз) и аэробную.
II
группа – дефосфорилирование, расщепление
АТФ и высвобождение энергии. III
группа – фосфорилирование, сопряженное
с процессом окисления.
Процесс окисления
глюкозы вначале происходит без участия
кислорода (анаэробным или гликолитическим
путем) до пировиноградной или молочной
кислоты.
Однако при этом
энергии выделяется лишь небольшое
количество. В дальнейшем эти кислоты
вовлекаются в процессы окисления,
которые протекают с участием кислорода,
т. е. являются аэробными. В результате
процесса окисления пировиноградной и
молочной кислоты, названной циклом
Кребса, образуется углекислый газ, вода
и большое количество энергии.
Образующаяся
энергия не выделяется в виде тепла, что
привело бы к перегреванию клеток и
гибели всего организма, а аккумулируется
в удобной для хранения и транспорта
форме в виде аденозинтрифосфорной
кислоты (АТФ). Синтез АТФ происходит из
АДФ и фосфорной кислоты и вследствие
этого называется фосфорилированием.
В здоровых клетках
фосфорилирование сопряжено с окислением.
При заболеваниях сопряженность может
разобщаться, поэтому субстрат окисляется,
а фосфорилирование не происходит, и
окисление переходит в тепло, а содержание
АТФ в клетках снижается. В результате
повышается температура и падает
функциональная активность клеток.
Итак,
основная функция митохондрий
заключается
в выработке практически
всей энергии клетки и происходит синтез
компонентов, необходимых для деятельности
самого органоида, ферментов «дыхательного
ансамбля», фосфолипидов и белков.
Еще одной стороной
деятельности митохондрий является их
участие в специфических синтезах,
например, в синтезе стероидных гормонов
и отдельных липидов. В ооцитах разных
животных образуются скопления желтка
в митохондриях, при этом они утрачивают
свою основную систему. Отработавшие
митохондрии могут накапливать также
продукты экскреции.
В
некоторых
случаях (печень, почки) митохондрии
способны аккумулировать вредные вещества
и яды, попадающие в клетку, изолируя их
от основной цитоплазмы и частично
блокируя вредное действие этих веществ.
Таким образом, митохондрии способны
брать на себя функции других органоидов
клетки, когда это требуется для
полноценного обеспечения того или иного
процесса в норме или в экстремальных
условиях.
Биогенез
митохондрий.
Митохондрии
представляют собой обновляющиеся
структуры с довольно кратким жизненным
циклом (в клетках печени крысы, например,
период полужизни митохондрий охватывает
около 10 дней). Митохондрии образуются
в результате роста и деления предшествующих
митохондрий. Деление их может происходить
тремя способами: перетяжкой, отпочковыванием
небольших участков и возникновением
дочерних митохондрий внутри материнской.
Делению (репродукции) митохондрий
предшествует репродукция собственной
генетической системы – митохондриальной
ДНК.
Итак,
согласно взглядам большинства
исследователей, образование митохондрий
происходит преимущественно путем
саморепродукции их de
novo.
Чего боится митохондрия?
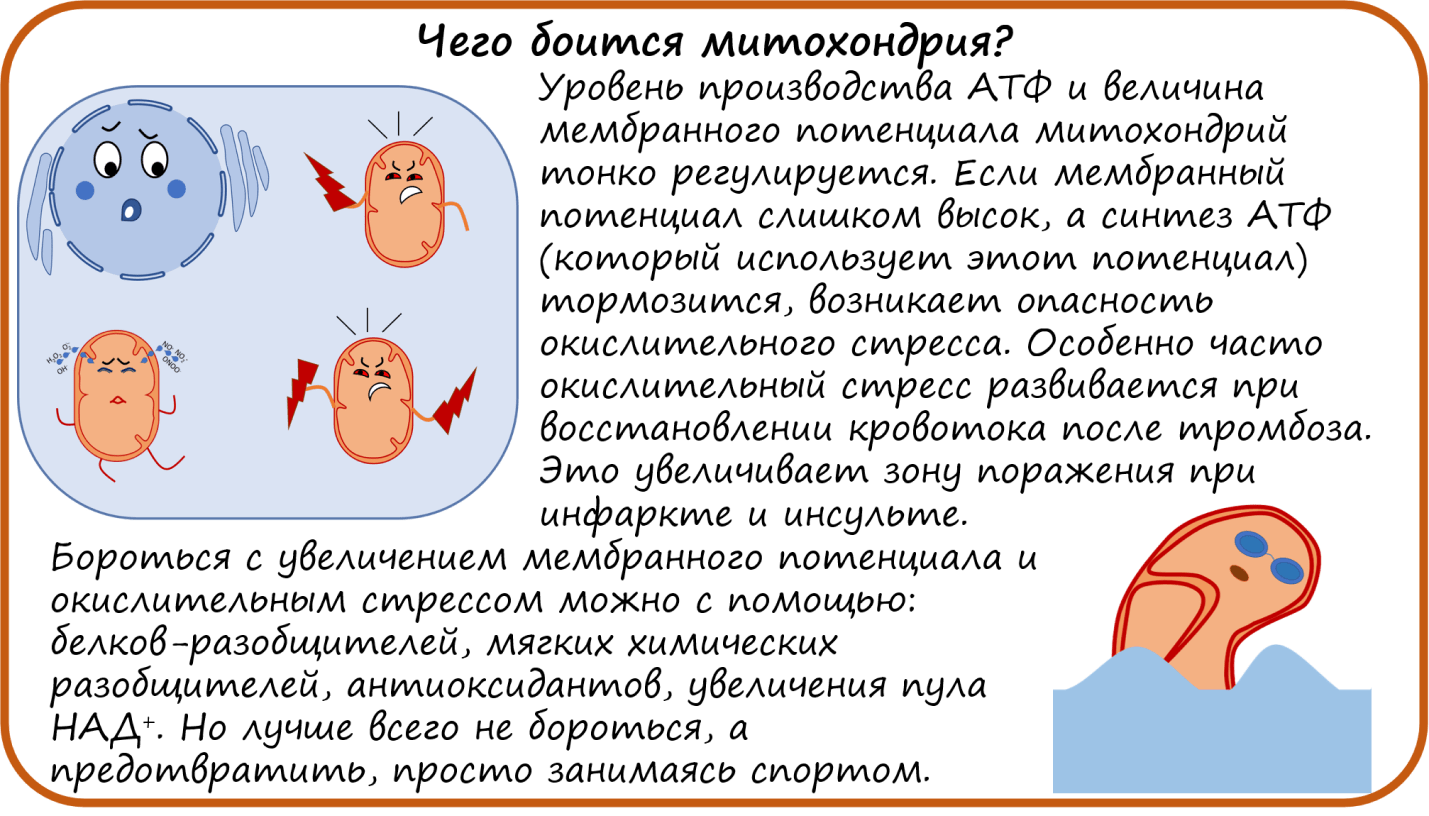
Итак, мы разобрались, как митохондрии производят АТФ. Причем количество АТФ должно очень тонко регулироваться клеткой. Если потребности в АТФ выросли, его надо произвести больше. Например, при активной мышечной или умственной работе требуется больше АТФ. И наоборот, если клетка в спокойном состоянии, то и не надо тратить ресурсы понапрасну. Но как митохондрия понимает, когда простимулировать работу ЭТЦ, а когда — притормозить?
Поддерживать работу ЭТЦ на определенном уровне важно еще и потому, что ЭТЦ обеспечивает постоянство мембранного потенциала. Если мембранный потенциал слишком низкий, в митохондрии нарушается транспорт веществ. Но и слишком большой потенциал — это тоже плохо. Это значит, что в межмембранном пространстве митохондрий накопилось слишком много протонов. Перенос каждого следующего протона оказывается сильно затруднен. А перенос электронов, как мы помним, сопровождается закачкой протонов в межмембранное пространство. Без движения протонов через мембрану не будет и движения электронов вдоль мембраны. Поэтому электроны оказываются на долгое время «заперты» в своих комплексах ЭТЦ. И там-то их может схватить кислород. В таком случае образуется не безобидная вода, а активные формы кислорода (АФК, или кислородные радикалы), которые разрушают все на своем пути. В первую очередь, это супероксид-анион радикал (О2*—) и перекись водорода (Н2О2). Накопление таких агрессивных соединений провоцирует окислительный стресс — губительное состояние для тканей.
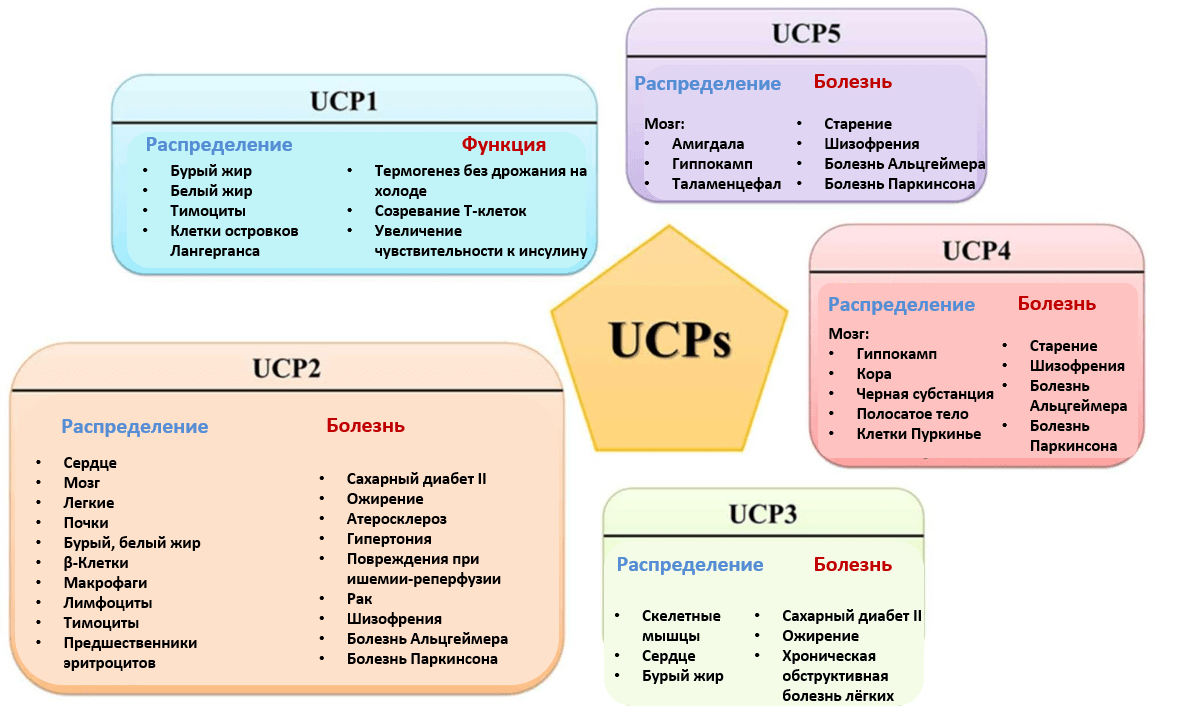
Рисунок 4. Семейство белков-разобщителей электронного транспорта и синтеза АТФ (UCPs — uncoupling proteins).
Другой способ снизить потенциал на мембране митохондрий — это увеличить пул НАД+. Как это работает? Комплекс I ЭТЦ окисляет НАДН (то есть отбирает у него электроны), в результате чего электроны поступают в ЭТЦ для создания мембранного потенциала и производства АТФ, а НАДН превращается в НАД+. Однако, комплекс I «чувствует» баланс НАД+/НАДН, и при нарушении баланса, в данном случае — увеличении количества НАД+, комплекс I перестает окислять НАДН. Без притока электронов с молекул НАДН протоны не закачиваются в межмембранное пространство, и мембранный потенциал митохондрий снижается.
В клетках существуют собственные антиоксидантные системы. Например, фермент супероксиддисмутаза преобразует опасный супероксидный анион радикал (О2*—) в кислород и пероксид водорода. Пероксид водорода тоже считается АФК, которая может вызвать окислительный стресс. Для его утилизации работает фермент каталаза, разлагающий пероксид водорода до воды и кислорода.
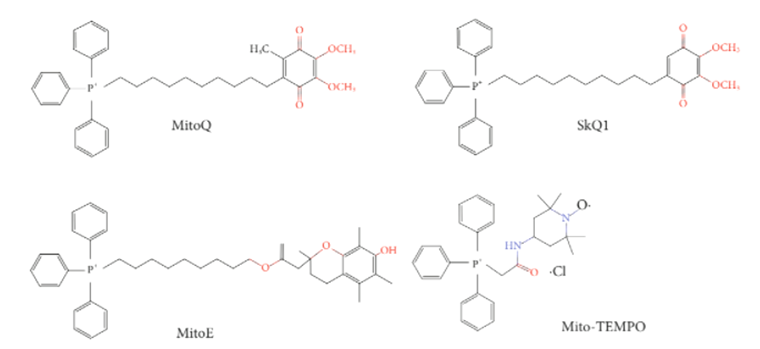
Рисунок 5. Антиоксиданты, соединенные с липофильными катионами.
Итак, мы рассмотрели, как устроена ЭТЦ митохондрий, как производится АТФ, зачем нам нужен кислород. Но в жизни все не так просто, как на картинке. Комплексы ЭТЦ могут еще и объединяться в суперкомплексы — так называемые респирасомы (рис. 6).
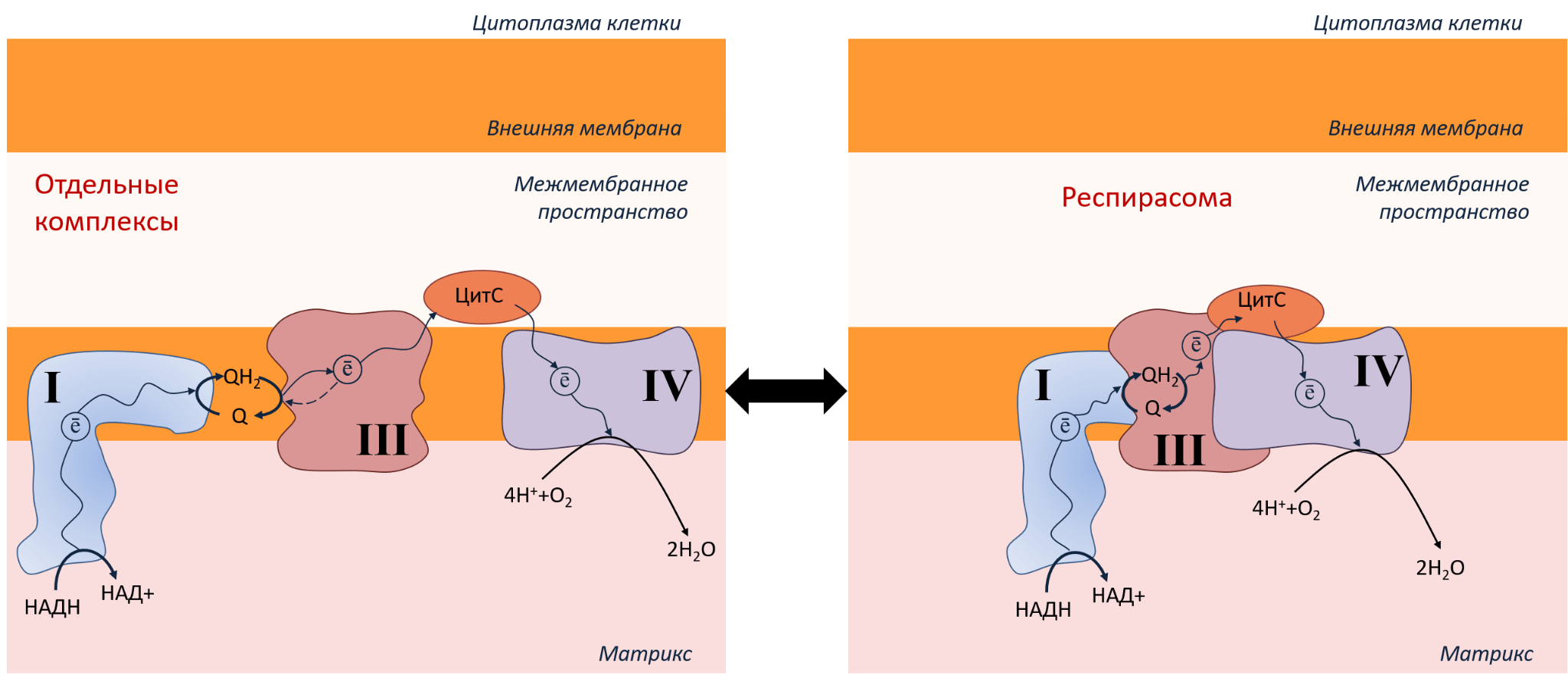
Рисунок 6. ЭТЦ в митохондриях может быть организована в виде отдельных комплексов (слева) или в виде респирасомы (справа). Переход из одного состояния в другое зависит от типов клеток и условий.
Респирасомы повышают эффективность переноса электрона по комплексам ЭТЦ, что ускоряет производство АТФ и снижает энергетические потери. Белки держатся внутри комплекса благодаря нескольким механизмам:
- за счет электростатических взаимодействий: если на поверхности одного комплекса расположены аминокислоты с положительным зарядом, а на поверхности другого комплекса — аминокислоты с отрицательным зарядом, то такие комплексы лучше удерживаются вместе;
- при участии особого липида, который есть только во внутренней мембране митохондрий — кардиолипина (несмотря на название, он есть не только в сердце, но и во всех митохондриях, а также у бактерий). Рассмотрим его роль подробнее.
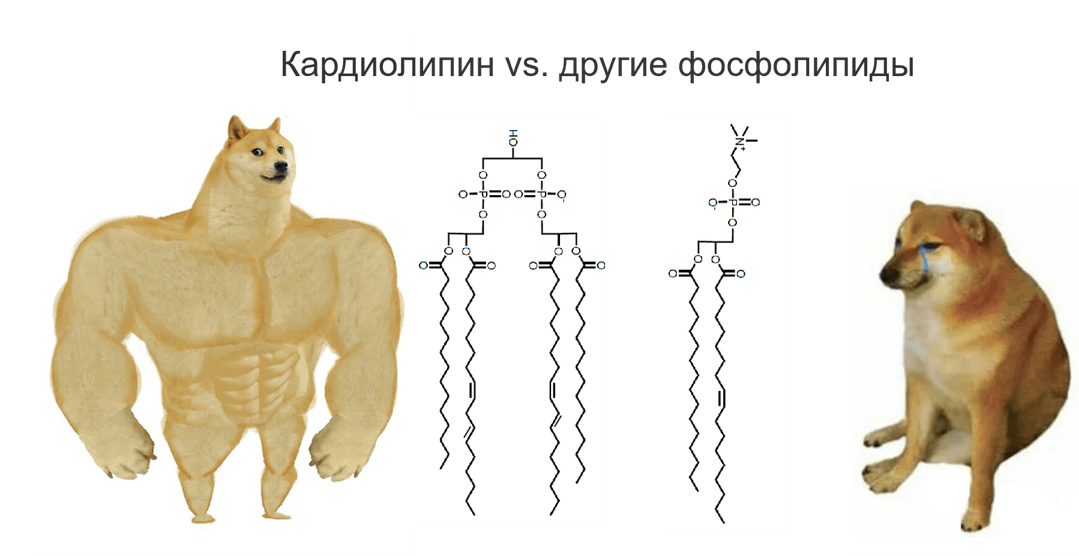
Рисунок 7. Принцип строения кардиолипина и других фосфолипидов.
Междисциплинарность — залог успеха
В 2010 году, когда интерес к нанотехнологиям был на высоте, академик Юрий Дмитриевич Третьяков, в то время декан факультета наук о материалах (ФНМ) МГУ, решил начать межфакультетское сотрудничество с биологами для проведения совместных исследований методом ГКР. В итоге сотрудничество было установлено с лабораторией биофизики клетки под руководством Г.В. Максимова. Так наша лаборатория познакомилась с заместителем декана ФНМ химиком-материаловедом член-корр. Е.А. Гудилиным. Совместно с группой Гудилина начались активные исследования по применению ГКР в биологии.
Дело в том, что ГКР — метод непростой и требует исключительно междисциплинарного подхода. Каждый биообъект уникален, и чтобы регистрировать от него сигнал ГКР, нужно разрабатывать уникальные наноструктуры, которые бы усиливали сигнал от нужного объекта, не повреждали его и выдерживали «атаку» физиологических многокомпонентных буферов.
Раз это удалось сделать для эритроцитов, то можно сделать и для митохондрий!
Как всё устроено
* — Окислительно-восстановительный потенциал — синоним редокс-потенциала (от англ. reduction — восстановление, oxidation — окисление).
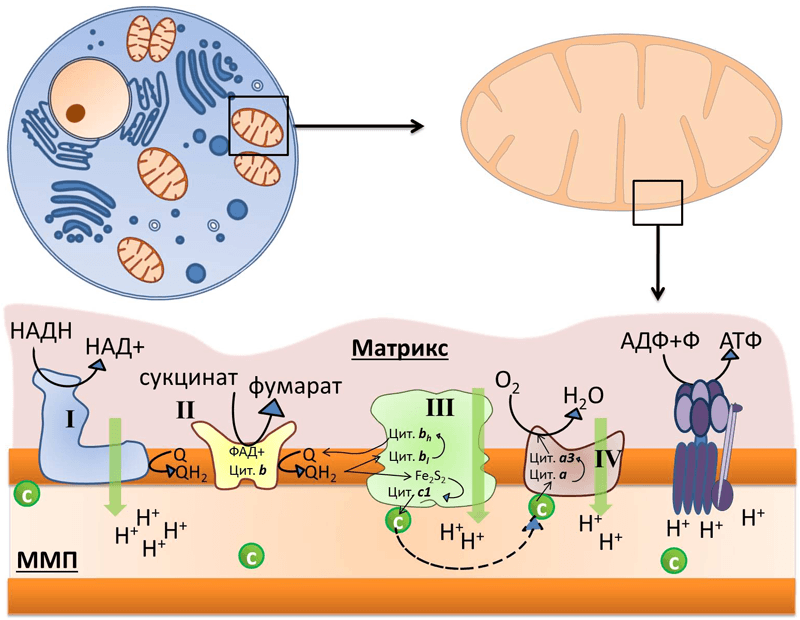
Рисунок 1. Во внутренней мембране митохондрий локализованы комплексы дыхательной цепи: I — НАДН-дегидрогеназа, II — сукцинат-дегидрогеназа, III — цитохром b–c1-комплекс, или цитохром с-редуктаза, IV — цитохром а–а3, или цитохромоксидаза, и фермент АТФ-синтаза. Q — убихинон, «с» в зелёном кружке — цитохром с. Чёрными стрелками показан транспорт электрона, пунктирной стрелкой — диффузия цитохрома с от комплекса III к IV, зелеными стрелками — перенос протона.
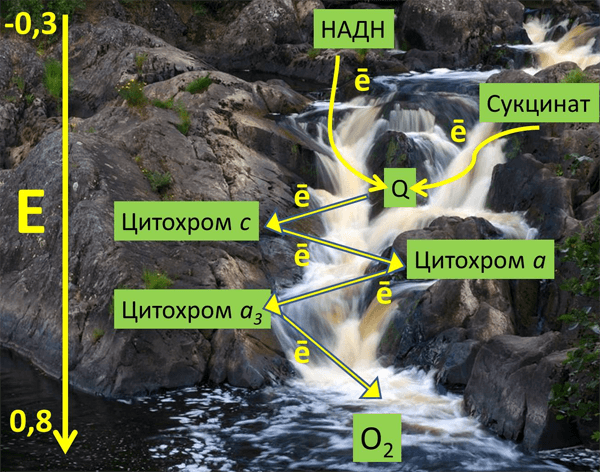
Рисунок 2. Метафорическое изображение среднеточечных редокс-потенциалов различных переносчиков ЭТЦ митохондрий в энергетической шкале (вольт) на фоне реки Тохмайоки в Карелии.
Как митохондрии управляют судьбой клетки?
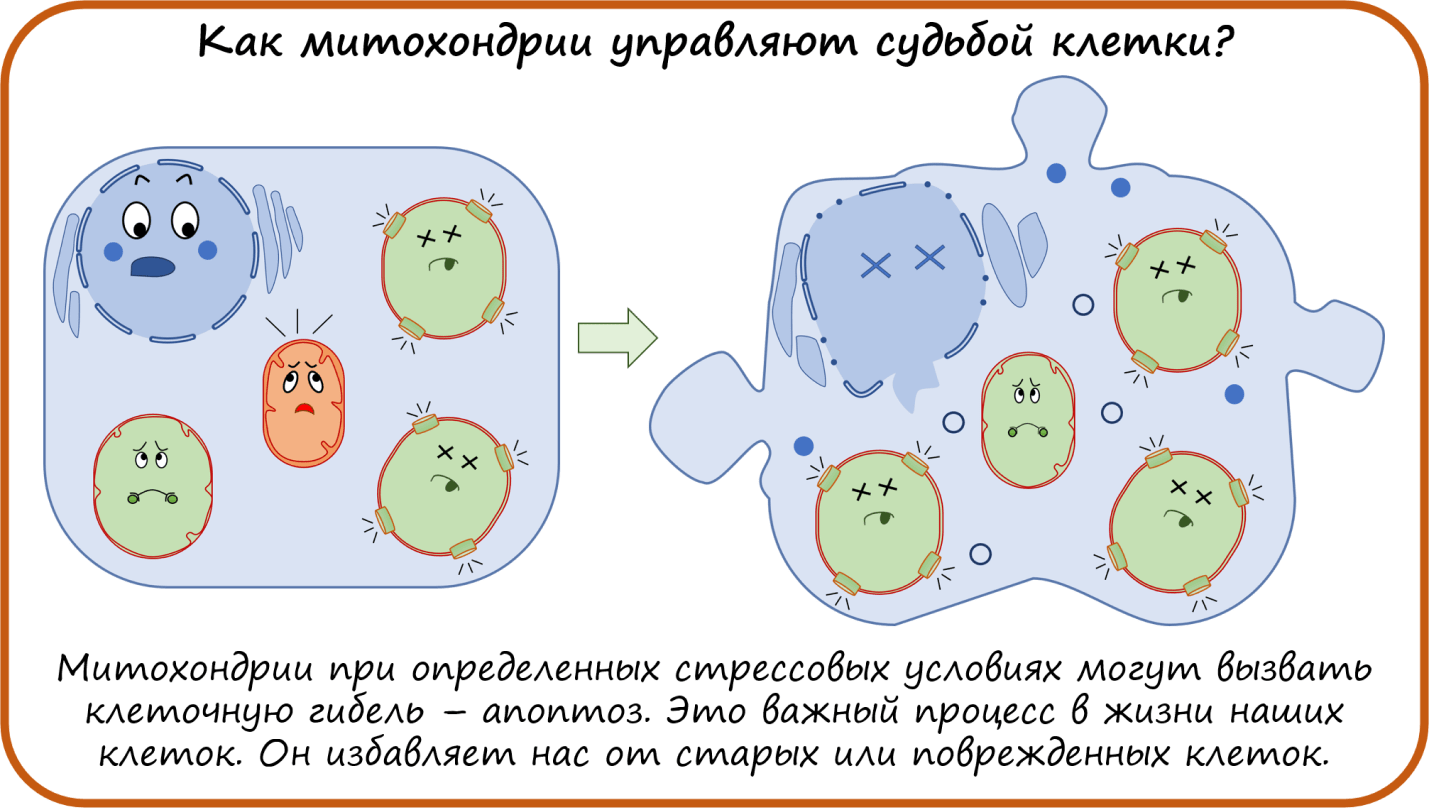
Вот мы и рассмотрели функции митохондрии как источника энергии и самонаводящейся ракеты при индукции апоптоза. Но кроме этого, митохондрии еще участвуют в синтезе белков, кофакторов, гормонов, нуклеотидов, что делает их еще более важными и ценными.
Этиология и патогенез митохондриальных заболеваний
Основными
причинами
митохондриальных заболеваний являются
мутации
митохондриальных генов, мутации генов
яДНК необходимых для работы митохондрий,
нарушение интергеномного взаимодействия,
которое может привести к возникновению
феномена деплеции — истощению числа
копий мтДНК, т. к. синтез мтДНК находится
под контролем яДНК.
Наследование
мутаций мтДНК имеет следующие особенности:
- Гетероплазмия
— наличие
в ооците мутантных и нормальных копий
мтДНК.
Поэтому сиблинги могут наследовать от
матери мутантную мтДНК,
но фенотипически отличаться. - Эффект «бутылочного
горлышка»
— уменьшение количества и неравномерное
распределение митохондрий при
формировании ооцитов. - Пороговый эффект
— для
проявления мутаций в энергозависимых
тканях необходима доля мутантных ДНК
выше 60–70 %, в менее энергозависимых —
выше 90 %. - Вариация доли
мутантных молекул в разных тканях.
Таблица
1 — Типы наследования митохондриальных
заболеваний (по С. Н. Иллариошкину,
2007)
Патогенез
митохондриальных болезней связан с
нарушением биохимических процессов,
происходящих в митохондриях.
Явление гетероплазмии
определяет существование в одной клетке
нормальных митохондрий и митохондрий
с нарушенной функцией. За счет первых
клетка может функционировать какое-то
время. Если продукция энергии в ней
падает ниже определенного порога, то
происходит компенсаторная пролиферация
всех митохондрий, включая дефектные. В
худшем положении оказываются клетки,
которые потребляют много энергии:
нейроны, мышечные волокна, кардиомиоциты.
Из-за утечки в
дыхательной цепи митохондрии постоянно
продуцируют свободные радикалы на
уровне 1–2 % поглощенного кислорода.
Количество продукции радикалов зависит
от мембранного потенциала митохондрий,
на изменения которого влияет состояние
АТФ-зависимых калиевых каналов
митохондрий. Открытие этих каналов
влечет за собой возрастание образования
свободных радикалов, повреждение других
белков митохондриальных мембран и
мтДНК. ДНК митохондрий не защищена
гистонами и хорошо доступна для радикалов,
что проявляется в изменении уровня
гетероплазмии. Принято считать, что
наличие 10 % митохондрий с измененной
ДНК не оказывает влияния на фенотип.
Соседние файлы в папке Печать
Немембранные органеллы. Рибосомы.
К
немембранным органеллам общего
назначения
относятся 1рибосомы,
2центриоли, 3микротрубочки, 4микрофиламенты,
промежуточные 5филаменты.
Рибосомы—
элементарные аппараты синтеза
белковых, полипептидных молекул
— обнаруживаются во всех клетках.
Рибосомы — это сложные рибонуклеопротеиды,
в состав которых входят
белки и молекулы РНК
примерно в равных весовых отношениях.
Размер функционирующей рибосомы
эукариотических клеток 25x20x20 нм. Такая
рибосома состоит из большой и малой
субъединиц.
Каждая из субъединиц построена из
рибонуклеопротеидного тяжа, где рРНК
взаимодействует с разными белками и
образует тело рибосомы.
Различают
единичные рибосомы и комплексные
рибосомы (полисомы).
Рибосомы
могут располагаться свободно в гиалоплазме
или быть связанными с ЭПС. В
малоспециализированных
и быстрорастущих клетках в основном
обнаруживаются свободные
рибосомы. В
специализированных
клетках рибосомы располагаются в составе
гранулярной
эндоплазматической сети.
Степень интенсивности синтетической
деятельности свободных рибосом меньше,
а образуемые белки используются в
основном на собственные нужды клетки.
Связанные рибосомы обеспечивают синтез
белков «на экспорт», т.е. на обеспечение
нужд организма. Содержание РНК и
соответственно степень белковых синтезов
коррелируют с интенсивностью базофилии
цитоплазмы.
Соседние файлы в предмете Гистология
Могут ли митохондрии передвигаться?
Долгожданный спектр ГКР митохондрий
Жалюзи опущены. Выключен свет. Комната погружена во мрак. Капля суспензии падает на серебряную подложку. Вспышка зелёного света. 30 секунд. Спектр. Тот самый долгожданный спектр ГКР от изолированных сердечных митохондрий был получен! Оставалось только понять, от каких именно структур в митохондриях исходит сигнал.
С учётом размеров компонентов митохондрий (рис. 7) и того, что усиление наблюдается на расстоянии нескольких нанометров от наноструктур, можно было предположить, что спектр ГКР митохондрий будет преимущественно спектром цитохрома с, так как этот цитохром наиболее близко подходит ко внешней мембране митохондрий и, соответственно, к наноструктурам. В то же время остальные цитохромы закреплены в комплексах внутренней мембраны.
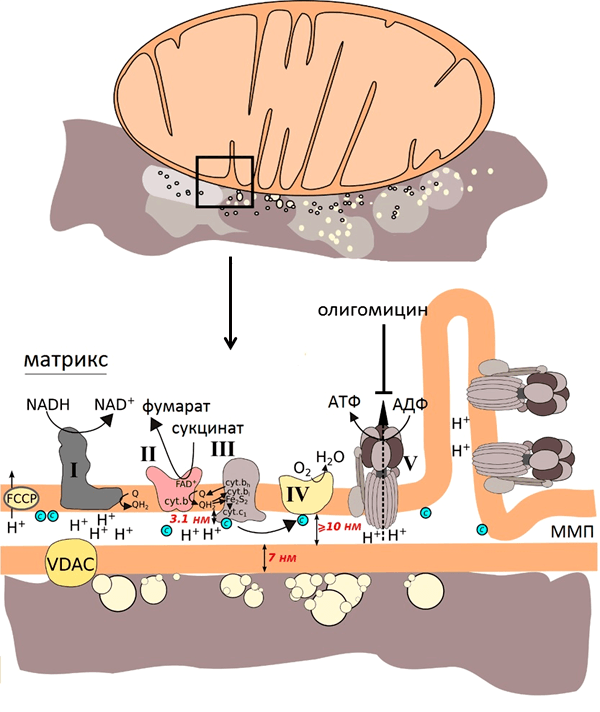
Это утверждение также подтверждалось при моделировании эффектов усиления КР серебряными наноструктурами, которые были использованы в работе. Группа наших коллег из лазерного центра Ганновера показала, что сложная морфология подложки со множеством углублений, в которые могут попадать митохондрии, позволило получать усиление на большом расстоянии (более 10 нм). А иерархическое устройство самих наноструктур увеличивало число мест контакта с мембраной митохондрий и, следовательно, число молекул цитохрома с, от которых можно было зарегистрировать сигнал ГКР. Если использовать те же наночастицы серебра, просто присоединенные к плоской подложке, то особого усиления не произойдет, что и подтверждалось в эксперименте (рис. 8).
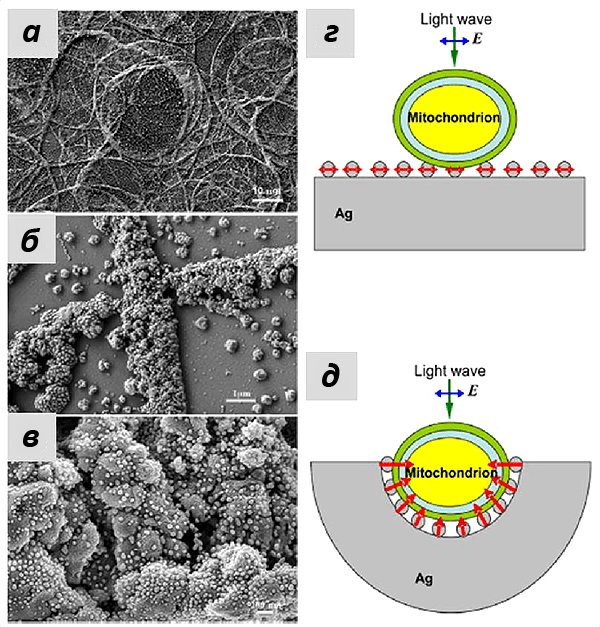
И действительно, полученный спектр ГКР митохондрий соответствовал спектрам цитохрома с! При использовании зелёного лазера в качестве возбуждающего света можно регистрировать сигнал только от цитохромов b и с, но не от цитохрома а, что облегчает задачу расшифровки спектров. Несмотря на схожесть структуры цитохромов типа b и с, они имеют ключевые пики на спектре, благодаря которым их нельзя перепутать (рис. 9). Таким образом, используемые наноструктуры давали усиление на достаточно большом расстоянии, чтобы зарегистрировать спектры от цитохрома с, но недостаточно большом, для того чтобы увидеть пики цитохромов b. И это как раз то, что нужно! Благодаря методу ГКР теперь можно исследовать селективно редокс-состояние и конформацию именно цитохрома с в живых функционирующих митохондриях.
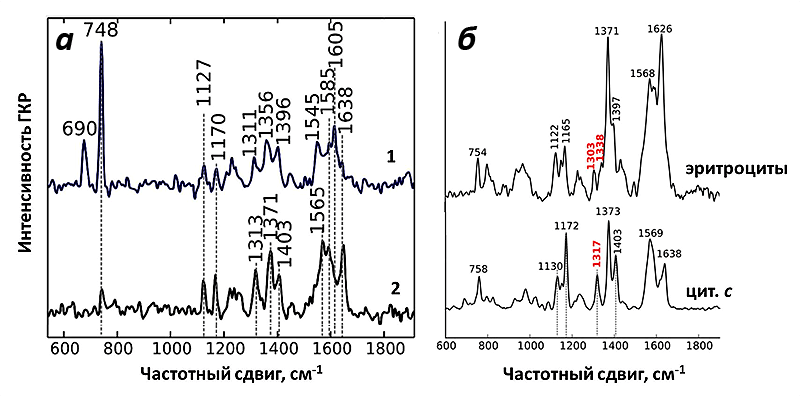
Как и ожидалось, спектры ГКР цитохрома с митохондрий оказались очень чувствительны к изменению его окислительно-восстановительного состояния. Для этого было исследовано два воздействия: внесение протонофора FCCP и ингибитора АТФ-синтазы олигомицина.
Таким образом, спектры ГКР митохондрий, являясь спектрами исключительно цитохрома с, оказались чувствительны к изменениям его конформации и редокс-состояния в процессе работы митохондрий.
Схема всей работы в одном ролике
К митохондриям от чистого сердца
Однако у цитохромов есть одна особенность — спектр КР интенсивен только для восстановленных их форм, тогда как спектр окисленных состояний обладает очень низкой интенсивностью. Поэтому основным критерием определения восстановленности цитохромов является интенсивность сигнала. Но если мы ориентируемся только на интенсивность, то можем потерять важную информацию об изменениях в исследуемой системе! Ведь интенсивность зависит и от количества молекул, и от выраженности тех или иных колебаний, и от других параметров. К тому же, для определения точного положения пиков большинство исследователей, имеющих дело с цитохромами, вынуждены искусственно восстанавливать образцы, что заведомо нарушает их нативность.
Усиливая сигнал
Идея пришла от О.В. Сосновцевой, профессора Копенгагенского университета, с которой наша лаборатория сотрудничает не первый год. Она даже предложила «подсобить» с митохондриями, так как с ними работали в соседней лаборатории, и они готовы были поделиться.
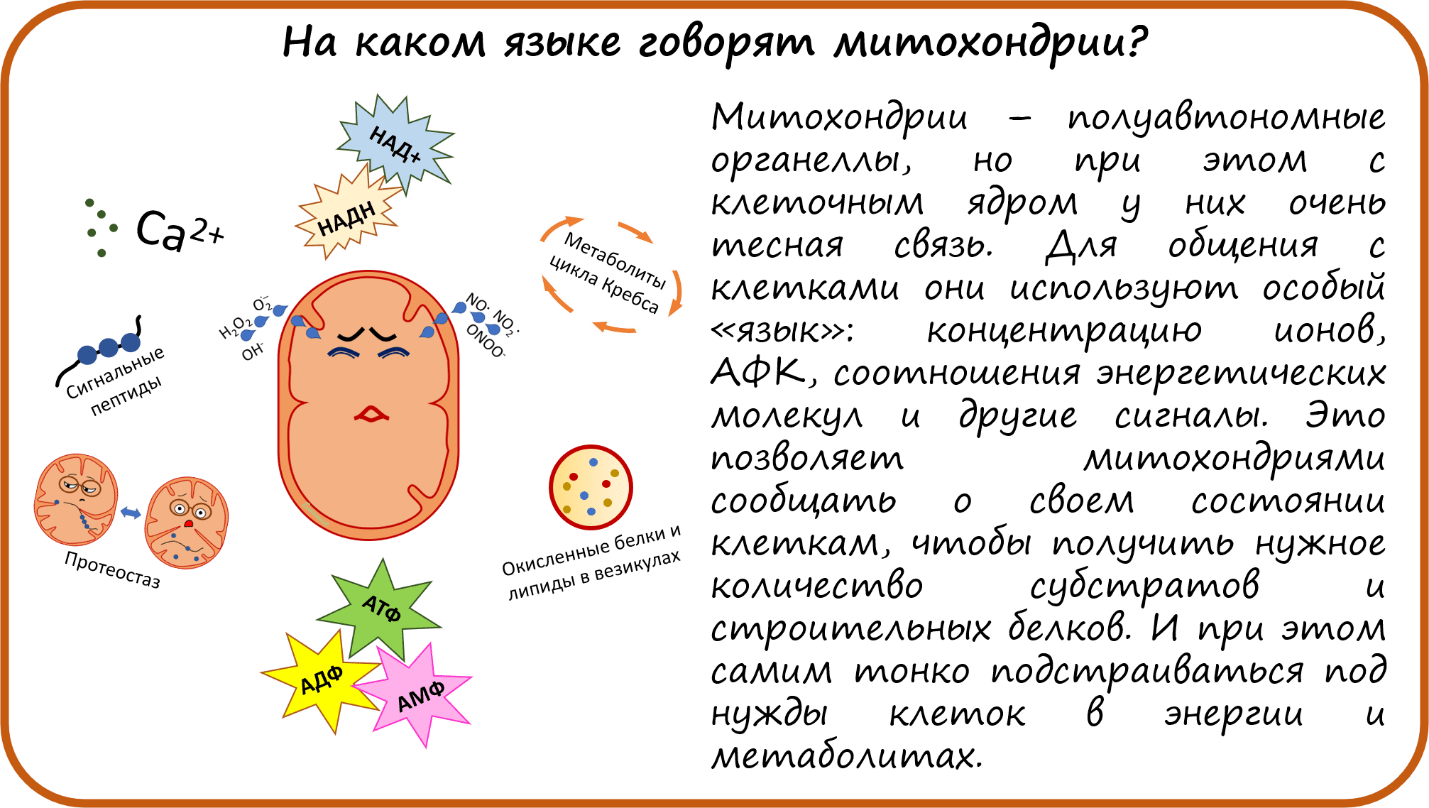
На каком языке говорят митохондрии?