
Гликоген
(С6Н10О5)n – резервный полисахарид, который
содержится в животных организмах. В
животных организмах гликоген локализируется
в печени (20%) и мышцах (4%).
Молекулы
гликогена имеют разветвленную структуру
и состоят из остатков альфа-D-глюкозы,
соединенных 1,4- и 1,6-гликозидными связями.
Гликоген
растворяется в горячей воде, с растворов
осаждается этиловым спиртом. Гликоген
устойчив в щелочной среде, а в кислой
среде при нагревании гидролизируется
с образованием сначала декстринов, а
потом глюкозы. В организме гликоген
является резервным полисахаридом,
который легко может превращаться в
глюкозу. При голодании и значительной
физической нагрузке содержание гликогена
в печени и мышцах значительно уменьшается.
Гликоген мышц и печени подвергается
постоянному расщеплению и синтезу.
Синтез и расщепление гликогена
регулируется нервно-гуморальными
системами (ЦНС, инсулином, адреналином).
Нарушение обмена гликогена наблюдается
при различных наследственных заболеваниях.
КАТАБОЛИЗМ И БИОСИНТЕЗ
ГЛИКОГЕНА. РЕГУЛЯЦИЯ ОБМЕНА ГЛИКОГЕНА. БИОСИНТЕЗ ГЛЮКОЗЫ – . МЕХАНИЗМЫ ГОРМОНАЛЬНОЙ
РЕГУЛЯЦИИ И ПАТОЛОГИЯ ОБМЕНА УГЛЕВОДОВ
СТРОЕНИЕ И ФУНКЦИИ ГЛИКОГЕНА

Рис. Структура гликогена. А.
Строение молекулы гликогена: 1 – остатки глюкозы, соединённые
α-1,4-гликозидной связью; 2 – остатки глюкозы, соединённые
α-1,6-гликозидной связью; 3 – нередуцирующие концевые мономеры; 4 –
редуцирующий концевой мономер. Б. Строение отдельного фрагмента молекулы
гликогена.
Таким образом, в молекуле гликогена
имеется только одна свободная аномерная ОН-группа и, следовательно, только один
восстанавливающий (редуцирующий) конец.
В клетках животных гликоген – основной
резервный полисахарид. При полимеризации глюкозы снижается растворимость
образующейся молекулы гликогена и, следовательно, её влияние на осмотическое
давление в клетке. Это обстоятельство объясняет, почему в клетке депонируется
гликоген, а не свободная глюкоза.
Гликоген хранится в цитозоле клетки в
форме гранул диаметром 10-40 нм. С гранулами связаны и некоторые ферменты,
участвующие в метаболизме гликогена, что облегчает их взаимодействие с субстратом.
Разветвлённая структура гликогена обусловливает большое количество концевых
мономеров, что способствует работе ферментов, отщепляющих или присоединяющих
мономеры при распаде или синтезе гликогена, так как эти ферменты могут
одновременно работать на нескольких ветвях молекулы. Гликоген депонируется
главным образом в печени и скелетных мышцах.
После приёма пищи, богатой углеводами,
запас гликогена в печени может составлять примерно 5% от её массы. В мышцах
запасается около 1% гликогена, однако масса мышечной ткани значительно больше и
поэтому общее количество гликогена в мышцах в 2 раза больше, чем в печени.
Гликоген может синтезироваться во многих клетках, например в нейронах,
макрофагах, клетках жировой ткани, но содержание его в этих тканях незначительно.
В организме может содержаться до 450 г
гликогена.
Распад гликогена
печени служит в основном для поддержания уровня глюкозы в крови в
пост-абсорбтивном периоде. Поэтому содержание гликогена в печени изменяется в
зависимости от ритма питания. При длительном голодании оно снижается почти до
нуля. Гликоген мышц служит резервом глюкозы – источника энергии при мышечном
сокращении. Мышечный гликоген не используется для поддержания уровня глюкозы в
крови. Как уже упоминалось ранее, в клетках мышц нет фермента
глюкозо-6-фосфа-тазы, и образование свободной глюкозы невозможно. Расход
гликогена в мышцах зависит в основном от физической нагрузки (рис.).
Рис.. Функции гликогена в печени
и мышцах.
Известно,
что фосфоролитический распад играет ключевую роль в мобилизации .
Рис.
Гормональная регуляция фосфоролитического
отщепления остатка
Фосфорилазы
переводят
(в частности, гликоген) из запасной формы в метаболически активную форму; в
присутствии фосфо-рилазы
распадается с образованием фосфорного эфира
(глюкозо-1-фосфата) без предварительного расщепления на более крупные обломки
.
В общей форме эту реакцию можно представить в следующем виде:
где
(С6Н10О5)n означает полисахаридную
цепь ,
а (С6Н10О5)n,– ту же цепь, но
укороченную на один глюкозный остаток. На рис. изображены процесс распада
до глюкозо-1-фосфата и участие в этом процессе цАМФ.
фосфорилаза существует в двух формах, одна из которых (фосфорилаза а) активна,
в то время как другая (фосфорилаза b) обычно неактивна. Обе формы могут
диссоциировать на субъединицы. Фосфорилаза b состоит из двух субъединиц, а
фосфорилаза а – из четырех. Превращение фосфо-рилазы b в фосфорилазу а
осуществляется
:
Катализируется
эта реакция ,
который называется киназой фосфорилазы b. Установлено, что эта киназа может
существовать как в активной, так и в неактивной форме. Неактивная киназа
фосфорилазы превращается в активную под влиянием
(киназа киназы фосфорилазы), и не просто ,
а цАМФ-зависимой .
Активная форма последней образуется при участии цАМФ, которая в свою очередь
образуется из
под действием
аденилатцик-лазы, стимулируемой, в частности,
и .
Увеличение содержания
в крови приводит в этой сложной цепи реакций к превращению фосфорилазы b в
фосфорилазу а и, следовательно, к освобождению
в виде глюкозо-1-фосфата из запасного
.
Обратное превращение фосфорилазы а в фосфорилазу b катализируется
(эта реакция практически необратима). Образовавшийся в результате
фосфоролитического распада
глюкозо-1-фосфат превращается под действием
в глюкозо-6-фосфат. Для осуществления данной реакции необходима
фосфорилированная форма ,
т.е. ее активная форма, которая образуется, как отмечалось, в присутствии
глюкозо-1,6-бисфосфата .
Образование
свободной
из глюкозо-6-фосфата в печени происходит под влиянием глюкозо-6-фосфатазы.
Данный
катализирует гидролитическое отщепление фосфата:
Рис.
Распад и синтез
Жирными
стрелками указан путь распада, тонкими – путь синтеза. Цифрами обозначены :
1 – фосфорилаза; 2 – фос-фоглюкомутаза; 3 – глюкозо-6-фосфатаза; 4 –
(глюкокиназа); 5 – глюко-зо-1-фосфат-уридилтрансфераза; 6 – глико-генсинтаза.
Заметим,
что фосфорилированная
в противоположность неэтерифицированной
не может легко диффундировать из .
Печень содержит гидролитический
глюкозо-6-фосфатазу, который и обеспечивает возможность быстрого выхода
из этого органа. В мышечной
глюкозо-6-фосфатаза практически отсутствует.На рис. отражены представления о путях распада и синтеза
в печени.Можно считать, что
сохранение постоянства
в крови является результатом одновременного протекания двух процессов:
поступления
в кровь из печени и потребления ее из крови ,
где она используется в первую очередь как энергетический материал.В
(в том числе в печени) распад
происходит двумя основными путями: анаэробным (при отсутствии кислорода) и
аэробным, для осуществления которого необходим .
Глюконеогенез
из неуглеводных продуктов. Такими продуктами или являются в первую очередь молочная и
пи-ровиноградная кислоты, так называемые гликогенные , гли-церол и ряд других соединений. Иными словами, может быть пируват или любое соединение,
превращающееся в процессе в пируват или один из промежуточных продуктов . У
позвоночных наиболее интенсивно печени и почек (в корковом веществе). Большинство
стадий представляет собой обращение реакции . Только 3 реакции (гексокиназная, фосфо-фруктокиназная и
пируваткиназная) необратимы, поэтому в процесс глю-конеогенеза на 3 этапах
используются другие . Рассмотрим путь синтеза
Образование
фосфоенолпирувата из пирувата. Синтез
фосфоенолпирувата осуществляется в несколько этапов. Первоначально пируват под
влиянием и при участии СО карбоксилируется с образованием оксалоацетата:
Затем
оксалоацетат в результате
и фосфорили-рования под влиянием
фосфоенолпируваткарбоксилазы превращается в фосфоенолпируват.
фосфатного остатка в реакции служит гуанозинтрифосфат (ГТФ):
Установлено,
что в процессе образования фосфоенолпирувата участвуют
цитозоля и .
Первый
этап синтеза протекает в .
Пируваткарбоксилаза, которая катализирует эту реакцию, является аллостерическим
митохондриальным .
В качестве аллостерического активатора данного
необходим ацетил-КоА.
непроницаема для образовавшегося оксалоацетата. Последний здесь же, в ,
восстанавливается в малат:
Реакция протекает при участии митохондриальной
НАД-зависимой .
В
отношение НАДН/НАД+ относительно велико, в связи с чем
внутримитохондриальный оксалоацетат легко восстанавливается в малат, который
легко выходит из
через митохондриальную .
В цитозоле отношение НАДН/НАД+ очень мало, и малат вновь окисляется
при участии цитоплазматической НАД-за-висимой :
Дальнейшее
превращение оксалоацетата в фосфоенолпируват происходит в цитозоле .
Превращение фруктозо-1,6-бисфосфата во
фруктозо-6-фосфат.
Фосфо-енолпируват, образовавшийся из пирувата, в результате ряда обратимых
реакций превращается во фруктозо-1,6-бисфосфат.
Далее следует фосфофруктокиназная реакция, которая необратима. идет в обход этой
эндергонической реакции. Превращение фруктозо-1,6-бис-фосфата во фруктозо-6-фосфат
катализируется специфической
Рис.
Образование фосфоенолпирувата из пирувата. 1 –
; 2 – малатдегидрогеназа (митохондриальная); 3
-малатдегидрогеназа (цитоплазматическая); 4 – фосфоенолпируваткарбоксикиназа.
Образование
глюкозы из глюкозо-6-фосфата. В последующей
обратимой стадии фруктозо-6-фосфат превращается в глюкозо-6-фосфат.
Последний может дефосфорилироваться (т.е. реакция идет в обход гексокиназной
реакции) под влиянием
На
рис. 10.7 представлены «обходные» реакции
при
из пирувата и лактата.
Регуляция
глюконеогенеза. Важным моментом в регуляции
глюконеоге-неза является реакция, катализируемая . Роль положительного аллостерического модулятора
этого выполняет ацетил-КоА. В отсутствие ацетил-КоА почти полностью лишен . Когда в накапливается митохондриальный ацетил-КоА, из пирувата усиливается. Известно, что ацетил-КоА
одновременно является отрицательным модулятором пируватдегидроге-назного
комплекса. Следовательно, накопление ацетил-КоА замедляет окислительное пирувата, что также способствует превращению
последнего в Другой
важный момент в регуляции
– реакция, катализируемая фруктозо-1,6-бисфосфатазой – ,
который ингибиру-ется .
Противоположное действие
оказывает на фосфофруктокиназу, т. е. для этого
он является аллостерическим активатором. При низкой
и высоком уровне
происходит стимуляция .
Напротив, когда величина отношения АТФ/АМФ мала, в
наблюдается расщепление .В
1980 г. группой бельгийских исследователей (Г. Херс и др.) в
печени был открыт фруктозо-2,6-бисфосфат, который является мощным регулятором
двух перечисленных :
Фруктозо-2,6-бисфосфат
активирует фосфофруктокиназу и ингибирует фруктозо-1,6-бисфосфатазу. Повышение
в
уровня фруктозо-2,6-бис-фосфата способствует усилению
и уменьшению скорости глю-конеогенеза. При снижении
фруктозо-2,6-бисфосфата отмечается обратная картина.Установлено,
что
фруктозо-2,6-бисфосфата происходит из фруктозо-6-фосфата при участии ,
а распадается он на фруктозо-6-фосфат и .
и распад фруктозо-2,6-бис-фосфата катализируется одним и тем же ,
т.е. данный
бифункционален, он обладает и фосфокиназной, и фосфатазной :
Показано
также, что бифункциональный
в свою очередь регулируется путем цАМФ-зависимого .
Фосфорилирова-ние приводит к увеличению фосфатазной
и снижению фосфо-киназной
бифункционального .
Этот механизм объясняет быстрое воздействие ,
в частности ,
на уровень фруктозо-2,6-бисфосфата в .
бифункционального
регулируется также некоторыми ,
среди которых наибольшее значение имеет гли-церол-3-фосфат. Действие
глицерол-3-фосфата на
по своей направленности аналогично эффекту, который наблюдается при его
фосфори-лировании с помощью цАМФ-зависимых .В настоящее время фруктозо-2,6-бисфосфат, помимо печени, обнаружен и в
других органах и
животных, а также у растений и .Показано, что
может регулироваться и непрямым путем, т.е. через изменение
,
непосредственно не участвующего в синтезе .
Так, установлено, что
пиру-ваткиназа существует в 2 формах – L и М. Форма L (от англ. liver – печень)
преобладает в ,
способных к .
Эта форма ингиби-руется избытком
и некоторыми ,
в частности ала-нином. М-форма (от англ. muscle – мышцы) такой регуляции не
подвержена. В условиях достаточного обеспечения
энергией происходит инги-бирование L-формы .
Как следствие ингибирования замедляется
и создаются условия, благоприятствующие глюконеогенезу.Наконец, интересно отметить, что между ,
интенсивно протекающим в мышечной
при ее активной деятельности, и глюко-неогенезом, особенно характерным для
печеночной ,
существует тесная взаимосвязь. При максимальной
мышц в результате усиления
образуется избыток , диффундирующей в кровь, в печени значительная ее часть
превращается в
(глюконеогенез). Такая
затем может быть использована как энергетический ,
необходимый для деятельности мышечной .
Взаимосвязь между процессами
в мышечной
и глюконеогенезом в печени может быть представлена в виде схемы:
Синтез гликогена (гликогенез)
Прежде всего подвергается при
участии
, а в печени –
и глюкокиназы. Далее глюкозо-6-фосфат под влиянием переходит
в глюкозо-1-фосфат:
Образовавшийся
глюкозо-1-фосфат уже непосредственно вовлекается в синтез . На первой
стадии синтеза глюкозо-1-фосфат вступает во взаимодействие с УТФ
(уридинтрифосфат), образуя уридиндифосфатглю-козу (УДФ-глюкоза) и пирофосфат.
Данная реакция катализируется
глюкозо-1-фосфат-уридилилтрансферазой (УДФГ-пирофосфорилаза):
Глюкозо-1-фосфат
+ УТФ УДФ-глюкоза + Пирофосфат.
Приводим структурную
формулу УДФ-глюкозы:
Образующийся УДФ затем
вновь фосфорилируется в УТФ за счет , и таким образом весь
цикл превращений глюкозо-1-фосфата начинается сначала. В целом образование
α-1,4-глюкозидной ветви («амилозной» ветви) можно
представить в виде следующей схемы:
При участии
последних происходит
ряда ,
в том числе гликогенсинтазы и фосфорилазы b – , участвующих в
обмене .
Фосфорилированный
гликогенсинтаза сам по себе малоактивен или полностью неактивен, но в
значительной мере активируется положительным модулятором глюкозо-6-фосфатом,
который увеличивает Vmax.
Эта форма гликогенсинтазы называется D-формой, или зависимой (dependent)
формой, поскольку ее
зависит от глюкозо-6-фосфата. Дефосфорилированная форма гликоген-синтазы,
называемая также I-формой, или независимой (independent) формой, активна и в
отсутствие глюкозо-6-фосфата.Таким
образом,
оказывает двойное действие на обмен :
ингибирует синтез
из УДФ-глюкозы, поскольку для проявления максимальной
D-формы гликогенсинтазы нужны очень высокие
глюкозо-6-фосфата, и ускоряет распад ,
так как способствует образованию активной фосфорилазы а. В целом суммарный
результат действия
состоит в ускорении превращения
в .
СИНТЕЗ И РАСПАД ГЛИКОГЕНА
– главная форма запасания у животных и
человека. Накапливается
главным образом в печени (до 6% от массы печени) и в скелетных мышцах, где его
содержание редко превышает 1%. Запасы в скелетных
мышцах ввиду значительно большей массы последних превышают его запасы в печени.
присутствует в цитозоле в форме гранул диаметром от 10 до 40 нм. На электронных
гликогеновые гранулы выглядят плотными. Установлено, что эти гранулы, кроме , содержат , катализирующие
синтез и распад .
Однако гликогеновые гранулы отличаются от мульти-ферментных комплексов
(например, от пируватдегидрогеназного комплекса). Степень структурной
организации гликогеновых гранул ниже, чем в мультиферментных комплексах.
Следует подчеркнуть, что синтез и распад в осуществляются
разными метаболическими путями. В частности, считалось, что
(фосфорилаза а) катализирует как распад , так и его
синтез, потому что в опытах in vitro было показано, что гликогенфосфорилазная
реакция обратима. Однако в дальнейшем было установлено, что в (in vivo) фосфорилаза
а катализирует только распад , синтез осуществляется
при участии совершенно другого . Оба эти процесса
(синтез и распад гликогена) регулируют содержание в крови и создают
резерв
для интенсивной мышечной работы.
Депонирование
и распад гликогена
– основная форма депонирования глюкозы в
клетках животных. У растений эту же функцию выполняет крахмал. В структурном отношении
гликоген, как и крахмал, представляет собой разветвленный полимер из глюкозы:
Однако
гликоген более разветвлен и компактен. Ветвление обеспечивает быстрое освобождение
при распаде гликогена большого количества концевых мономеров. Синтез и распад
гликогена не являются обращением друг в друга, эти процессы происходят разными
путями:
Биосинтез гликогена – гликогенез показан на рисунке:
Гликоген синтезируется в период пищеварения (в течение 1-2 часов
после приема углеводной пищи). Гликогенез особенно интенсивно протекает в
печени и скелетных мышцах. В начальных реакциях образуется UDF-глюкоза (реакция
3), которая является активированной формой глюкозы, непосредственно
включающейся в реакцию полимеризации (реакция 4). Эта последняя реакция
катализируется гликогенсинтазой, которая присоединяет глюкозу к
олигосахариду или к уже имеющейся в клетке молекуле гликогена, наращивая цепь
новыми мономерами. Для подготовки и включения в растущую полисахаридную цепь
требуется энергия 1 моль АТР и 1 моль UTP. Ветвление
полисахаридной цепи происходит при участии фермента амило- a -1,4– a
-1,6-гликозил-трансферазы путем разрыва одной a -1,4-связи и переноса
олигосахаридного остатка от конца растущей цепи к ее середине с образованием в
этом месте a -1,6-гликозидной связи . Молекула гликогена содержит до 1 млн.
остатков глюкозы, следовательно, на синтез расходуется значительное количество
энергии. Необходимость превращения глюкозы в гликоген связана с тем, что накопление
значительного количества глюкозы в клетке привело бы к повышению осмотического
давления, так как глюкоза хорошо растворимое вещество. Напротив, гликоген
содержится в клетке в виде гранул, и мало растворим. Распад гликогена –
гликогенолиз – происходит в период между приемами пищи.
Освобождение
глюкозы в виде глюкозо-1-фосфата ( ) происходит в
результате фосфоролиза, катализируемого фосфорилазой. Фермент отщепляет
концевые остатки один за другим, укорачивая цепи гликогена. Однако этот фермент
расщепляет только a -1,4 гликозидные связи. Связи в точке ветвления
гидролизуются ферментом , который отщепляет мономер глюкозы в
свободном виде:
Механизм действия фосфорилазы гликогена
Особенности метаболизма гликогена в печени и мышцах
Включение глюкозы
в метаболизм начинается с образования фосфоэфира – глюкозо-6-фосфата. В
клетках мышц и других органах эту реакцию катализирует фермент гексокиназа,
его Км менее 0,1 ммоль/л. В клетках печени эту же реакцию
катализирует глюкокиназа, значение Км которой примерно 10
ммоль/л. Это значит, что насыщение глюкокиназы происходит только при высокой
концентрации глюкозы. Различия в свойствах ферментов объясняют, почему в период
пищеварения глюкоза задерживается в основном в печени. Глюкокиназа при высокой
концентрации глюкозы в этот период максимально активна. Напротив, гексокиназа,
обладая большим сродством к глюкозе, способна выхватывать ее из общего
кровотока, где концентрация глюкозы ниже.
Обмен гликогена в печени и мышцах
Физиологическое значение гликогенолиза в печени и в мышцах
различно.
Мышечный гликоген является источником глюкозы для самой клетки. Гликоген печени
используется главным образом для поддержания физиологической концентрации
глюкозы в крови. Различия обусловлены тем, что в клетке печени присутствует
фермент глюкозо-6-фосфатаза, катализирующая отщепление фосфатной
группы и образование свободной глюкозы, после чего глюкоза поступает в
кровоток. В клетках мышц нет этого фермента, и распад гликогена
идет только до образования глюкозо-6-фосфата, который затем используется в
клетке.
СИНТЕЗ ГЛИКОГЕНА (ГЛИКОГЕНОГЕНЕЗ)
Гликоген синтезируется в период
пищеварения (через 1-2 ч после приёма углеводной пищи). Следует отметить, что
синтез гликогена из глюкозы (рис.), как и любой анаболический процесс, является
эндергоническим, т.е. требующим затрат энергии.
Глюкоза, поступающая в клетку,
фосфори-лируется при участии АТФ (реакция 1). Затем глюкозо-6-фосфат в ходе
обратимой реакции превращается в глюкозо-1-фосфат (реакция 2) под действием
фермента фосфоглюкомутазы. Глюкозо-1-фосфат по термодинамическому состоянию мог
бы служить субстратом для синтеза гликогена. Но в силу обратимости реакции
глюкозо-6-фосфат ↔ глюкозо-1-фосфат синтез гликогена из глюкозо-1-фосфата
и его распад оказались бы также обратимыми и поэтому неконтролируемыми. Чтобы
синтез гликогена был термодинамически необратимым, необходима дополнительная
стадия образования уридинди-фосфатглюкозы из УТФ и глюкозо-1-фосфата (реакция
3). Фермент, катализирующий эту реакцию, назван по обратной реакции:
УДФ-глюкопирофосфорилаза. Однако в клетке обратная реакция не протекает, потому
что образовавшийся в ходе прямой реакции пирофосфат очень быстро расщепляется
пирофосфатазой на 2 молекулы фосфата (рис.).
Рис. Образование УДФ-глюкозы.
Реакция образования УДФ-глюкозы
обусловливает необратимость всей серии реакций, протекающих при синтезе
гликогена. Этим же объясняется невозможность протекания распада гликогена путём
простого обращения процесса его синтеза.
Образованная УДФ-глюкоза далее
используется как донор остатка глюкозы при синтезе гликогена. Эту реакцию
катализирует фермент гликогенсинтаза (глюкозилтрансфераза). Поскольку в данной
реакции не используется АТФ, фермент называют синтазой, а не синтетазой.
Нуклеотидная часть УДФ-глюкозы играет существенную роль в действии гликоген
синтазы, выполняя функцию «рукоятки», при помощи которой фермент располагает
глюкозу в полисахаридной цепи в нужном положении. Кроме того, нуклеотидная
часть УДФ-глюкозы, по-видимому, необходима для узнавания субстрата при
катализе.
Так как гликоген в клетке никогда не
расщепляется полностью, синтез гликогена осуществляется путём удлинения уже
имеющейся молекулы полисахарида, называемой «затравка», или «праймер». К
«затравке» последовательно присоединяются молекулы глюкозы. Строением молекулы
«затравки» как бы предопределяется тип связи, который возникает в реакции
трансгликозилирования. Таким образом, синтезируется полисахарид, аналогичный по
строению с «затравочным». В состав «затравки» может входить белок гликогенин, в
котором к ОН-группе одного из тирозиновых остатков присоединена олигосахаридная
цепочка (примерно 8 остатков глюкозы). Глюкозные остатки переносятся
гликогенсинтазой на не-редуцирующий конец олигосахаридной цепочки и связываются
а-1,4-гликозидными связями. По окончании синтеза гликогенин остаётся включённым
в гранулу гликогена.
РАСПАД ГЛИКОГЕНА (ГЛИКОГЕНОЛИЗ)
Распад гликогена
или его мобилизация происходят в ответ на повышение потребности организма в
глюкозе. Гликоген печени распадается в основном в интервалах между приёмами
пищи, кроме того, этот процесс в печени и мышцах ускоряется во время физической
работы.
Распад гликогена (рис.) происходит путём
последовательного отщепления остатков глюкозы в виде глюкозо-1-фосфата.
Рис. Распад гликогена. В рамке
– фрагмент гликогена с точкой ветвления. Закрашенный кружок – глюкозный
остаток, связанный α-1,6-гликозидной связью; светлые и заштрихованные
кружки – глю-козные остатки в линейных участках и боковых ветвях, связанные
а-1,4-гликозидной связью. 1 – гликоген-фосфорилаза; 2 –
олигосахаридтрансфераза; 3 – α-1,6-глюкозидаза.
Гликозидная
связь расщепляется с использованием неорганического фосфата, поэтому процесс
называется фосфоролизом, а фермент гликогенфосфорилазой.
Так же как и
синтез, расщепление гликогена начинается с нередуцирующего конца
поли-сахаридной цепи. При этом наличие разветвлённой структуры гликогена
облегчает быстрое высвобождение глюкозных остатков, так как чем больше концов
имеет молекула гликогена, тем больше молекул гликогенфосфорилазы могут
действовать одновременно.
Гликогенфосфорилаза
расщепляет только α-1,4-гликозидные связи (реакция 1). Последовательное
отщепление глюкозных остатков прекращается, когда до точки ветвления остаётся 4
мономера. Подобная особенность в действии гликогенфосфорилазы обусловлена
размером и строением её активного центра.
Дальнейший
распад гликогена требует участия двух других ферментов. Сначала три оставшихся
до точки ветвления глюкозных остатка переносятся при участии олигосахаридтрансферазы
(реакция 2) на нередуцирующий конец соседней цепи, удлиняя её и таким образом
создавая условия для действия фосфорилазы. Оставшийся в точке ветвления
глюкозный остаток гидроли-тически отщепляется с помощью а-1,6-глюко-зидазы в
виде свободной глюкозы (реакция 3), после чего неразветвлённый участок
гликогена может вновь атаковаться фосфорилазой.
Считают, что перенос трёх остатков
глюкозы и удаление мономера из точки ветвления (реакции 2 и 3) катализирует
один и тот же фермент, который обладает двумя разными ферментативными
активностями – трансферазной и гликозидазной. Его называют «деветвящим»
ферментом (от англ. debranching enzyme).
Продукт действия гликогенфосфорилазы –
глюкозо-1-фосфат – затем изомеризуется в глю-козо-6-фосфат фосфоглюкомутазой.
Далее глю-козо-6-фосфат включается в процесс катаболизма или другие
метаболические пути. В печени (но не в мышцах) глюкозо-6-фосфат может
гидролизо-ваться с образованием глюкозы, которая выделяется в кровь. Эту
реакцию катализирует фермент глюкозо-6-фосфатаза. Реакция протекает в просвете
ЭР, куда с помощью специального белка транспортируется глюкозо-6-фосфат.
Фермент локализован на мембране ЭР таким образом, что его активный центр
обращён в просвет ЭР. Продукты гидролиза (глюкоза и неорганический фосфат)
возвращаются в цитоплазму также с помощью транспортных систем.
БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ОБМЕНА
ГЛИКОГЕНА В ПЕЧЕНИ И МЫШЦАХ
На рисунке приведена общая схема синтеза
и распада гликогена и регуляция этих процессов гормонами.
Сравнение этих
процессов позволяет сделать следующие выводы:
• синтез и
распад гликогена протекают по разным метаболическими путям;
• печень
запасает глюкозу в виде гликогена не столько для собственных нужд, сколько для
поддержания постоянной концентрации глюкозы в крови, и, следовательно,
обеспечивает поступление глюкозы в другие ткани.
Присутствие в
печени глюкозо-6-фосфатазы обусловливает эту главную функцию печени в обмене
гликогена;
• функция
мышечного гликогена заключается в освобождении глюкозо-6-фосфата, потребляемого
в самой мышце для окисления и использования энергии;
• синтез
гликогена – процесс эндергоничес-кий. Так на включение одного остатка глюкозы в
полисахаридную цепь используется 1 моль АТФ и 1 моль УТФ;
• распад
гликогена до глюкозо-6-фосфата не требует энергии;
• необратимость
процессов синтеза и распада гликогена обеспечивается их регуляцией.
егуляция метаболизма гликогена
Процессы накопления глюкозы в виде
гликогена и его распада должны быть согласованы с потребностями организма в
глюкозе как источнике энергии. Одновременное протекание этих метаболических
путей невозможно, так как в этом случае образуется «холостой» цикл,
существование которого приводит только к бесполезной трате АТФ.
Изменение направления процессов в
метаболизме гликогена обеспечивают регуляторные механизмы, в которых участвуют
гормоны. Переключение процессов синтеза и мобилизации гликогена происходит при
смене абсорбтивного периода на постабсорбтивный или состояния покоя организма
на режим физической работы. В переключении этих метаболических путей в печени
участвуют гормоны инсулин, глюкагон и адреналин, а в мышцах – инсулин и адреналин.
арактеристика гормонов, регулирующих
обмен гликогена
Первичным
сигналом для синтеза и секреции инсулина и глюкагона является изменение уровня
глюкозы в крови. В норме концентрация глюкозы в крови соответствует 3,3-5,5
ммоль/л (60-100 мг/дл).
белковый
гормон, синтезируется и секретируется в кровь β-клетками островков
Лангерханса поджелудочной железы. β-клетки чувствительны к изменениям
содержания глюкозы в крови и секретируют инсулин в ответ на повышение её
содержания после приёма пищи. Транспортный белок (ГЛЮТ-2), обеспечивающий
поступление глюкозы в β-клетки, отличается низким сродством к ней.
Следовательно, этот белок транспортирует глюкозу в клетку поджелудочной железы
лишь после того, как её содержание в крови будет выше нормального уровня (более
5,5 ммоль/л).
В β-клетках
глюкоза фосфорилируется глюкокиназой, имеющей также высокую Кm для
глюкозы – 12 ммоль/л. Скорость фосфорилиро-вания глюкозы глюкокиназой в
β-клетках прямо пропорциональна её концентрации в крови.
Синтез инсулина регулируется глюкозой.
Глюкоза (или её метаболиты), по-видимому, непосредственно участвуют в регуляции
экспрессии гена инсулина. Секреция инсулина и глюкагона также регулируется
глюкозой, которая стимулирует секрецию инсулина из β-клеток и подавляет
секрецию глюкагона из α-клеток. Кроме того, сам инсулин снижает секрецию
глюкагона (см. раздел 11).
–
«гормон голода», вырабатываемый α-клетками поджелудочной железы в ответ на
снижение уровня глюкозы в крови. По химической природе глюкагон – пептид.
выделяется
из клеток мозгового вещества надпочечников в ответ на сигналы нервной системы,
идущие из мозга при возникновении экстремальных ситуаций (например, бегство или
борьба), требующих внезапной мышечной деятельности. Адреналин является сигналом
«тревоги». Он должен мгновенно обеспечить мышцы и мозг источником энергии.
егуляция активности гликогенфосфорилазы
и гликогенсинтазы
Поскольку синтез
и распад гликогена протекают по различным метаболическим путям, эти процессы
могут контролироваться реципрокно. Влияние гормонов на синтез и распад
гликогена осуществляется путём изменения в противоположных направлениях
активности двух ключевых ферментов: гликогенсинтазы и гликогенфос-форилазы с
помощью их фосфорилирования и дефосфорилирования (рис.).
Рис. Изменение активности
гликогенфосфорилазы и гликогенсинтазы. Кружками
обозначены молекулы фермента: активные – чёрные, неактивные – белые.
ФП-фосфатаза (ГР) – фосфопротеинфосфатаза гранул гликогена.
Гликогенфосфорилаза
существует в 2 формах: 1) фосфорилированная – активная (форма а); 2)
дефосфорилированная – неактивная (форма в). Фосфорилирование осуществляется
путём переноса фосфатного остатка с АТФ на гидроксильную группу одного из
сериновых остатков фермента. Следствие этого – конформационные изменения
молекулы фермента и его активация.
Взаимопревращения 2 форм
гликогенфосфорилазы обеспечиваются действием ферментов киназы фосфорилазы и
фосфопротеинфосфатазы (фермент, структурно связанный с моле кулами гликогена).
В свою очередь, активность киназы фосфорилазы и фосфопротеинфосфатазы также
регулируется путём фосфорилирования и дефосфорилирования.
Активация киназы фосфорилазы происходит
под действием протеинкиназы А – ПКА (цАМФ-зависимой). цАМФ сначала активирует
про-теинкиназу А, которая фосфорилирует киназу фосфорилазы, переводя её в
активное состояние, а та, в свою очередь, фосфорилирует глико-генфосфорилазу.
Синтез цАМФ стимулируется адреналином и глюкагоном.
Активация фосфопротеинфосфатазы
происходит в результате реакции фосфорилирования, катализируемой специфической
протеинкиназой, которая, в свою очередь, активируется инсулином посредством
каскада реакций с участием Ras-белка, а также других белков и ферментов
(сигнальный Ras-путь). Активируемая инсулином протеинкиназа фосфорилирует и тем
самым активирует фосфопротеинфосфатазу. Активная фосфопротеинфосфатаза
дефос-форилирует и, следовательно, инактивирует киназу фосфорилазы и
гликогенфосфорилазу (рис.).
Рис. Влияние инсулина на
активность гликогенсинтазы и киназы фосфорилазы. ФП-фосфатаза
(ГР) фосфопротеинфосфатаза гранул гликогена. ПК (рр90S6) – протеинкиназа,
активируемая инсулином.
Активность
гликогенсинтазы также изменяется в результате фосфорилирования и
дефосфори-лирования. Однако есть существенные различия в регуляции
гликоген-фосфорилазы и гликогенсинтазы:
•фосфорилирование
гликогенсинтазы катализирует ПК А и вызывает её инактивацию;
•дефосфорилирование
гликогенсинтазы под действием фосфопротеинфосфатазы, наоборот, её активирует.
РЕГУЛЯЦИЯ МЕТАБОЛИЗМА ГЛИКОГЕНА В ПЕЧЕНИ
Как уже отмечалось, первичный сигнал для
синтеза инсулина и глюкагона – изменение концентрации глюкозы в крови. Инсулин
и глюкагон постоянно присутствуют в крови, но при смене абсорбтивного периода
на пос-табсорбтивный изменяется их относительная концентрация, что является
главным фактором, переключающим метаболизм гликогена в печени. Отношение
концентрации инсулина в крови к концентрации глюкагона называют
«ин-сулин-глюкагоновый индекс». В постабсорбтивном периоде инсулин-глюкагоновый
индекс снижается, и решающее значение в регуляции концентрации глюкозы в крови
приобретает концентрация глюкагона.
Глюкагон для
гепатоцитов служит внешним сигналом о необходимости выделения в кровь глюкозы
за счёт распада гликогена (гликогенолиза) или синтеза глюкозы из других веществ
– глюконеогенеза (этот процесс будет изложен позднее). Гормон связывается с
рецептором на плазматической мембране и активирует при посредничестве G-белка
аденилатциклазу, которая катализирует образование цАМФ из АТФ. Далее следует
каскад реакций, приводящий в печени к активации гликогенфос-форилазы и
ингибированию гликогенсинтазы (рис.).
Рис. Регуляция синтеза и распада
гликогена в печени глюкагоном и адреналином. 1 – глюкагон и
адреналин взаимодействуют со специфическими мембранными рецепторами. Комплекс
гормон-рецептор влияет на конформацию G-белка, вызывая диссоциацию его на
протомеры и замену в a-субъединице ГДФ на ГТФ; 2 – α-субъединица,
связанная с ГТФ, активирует аденилатциклазу, катализирующую синтез цАМФ из АТФ;
3 – в присутствии цАМФ протеинкиназа А (цАМФ-зависимая) обратимо
диссоциирует, освобождая обладающие каталитической активностью субъединицы С; 4
– протеинкиназа А фосфорилирует и активирует киназу фосфорилазы; 5 – киназа
фосфорилазы фосфорилирует гликогенфосфорилазу, переводя её в активную форму; 6
– протеинкиназа А фосфорилирует также гликогенсинтазу, переводя её в неактивное
состояние; 7 – в результате ингибирования гликогенсинтазы и активации
гликогенфосфорилазы гликоген включается в процесс распада; 8 – фосфодиэстераза
катализирует распад цАМФ и тем самым прерывает действие гормонального сигнала.
Комплекс а-субъединица-ГТФ затем распадается, а-, β- и γ-субъединицы
G-белка реассоциируются.
Этот механизм приводит к высвобождению
из гликогена глюкозо-1-фосфата, который превращается в глюкозо-6-фосфат. Затем
под влиянием глюкозо-6-фосфатазы образуется свободная глюкоза, способная выйти
из клетки в кровь. Таким образом, глюкагон в печени, стимулируя распад
гликогена, способствует поддержанию глюкозы в крови на постоянном уровне.
Адреналин
стимулирует выведение глюкозы из печени в кровь, для того чтобы снабдить ткани
(в основном мозг и мышцы) «топливом» в экстремальной ситуации. Эффект
адреналина в печени обусловлен фосфорилированием (и активацией)
гликогенфосфорилазы. Адреналин имеет сходный с глюкагоном механизм действия. Но
возможно включение и другой эффекторной системы передачи сигнала в клетку
печени (рис.).
Рис. Регуляция синтеза и распада
гликогена в печени адреналином и Са2+. ФИФ2
– фосфатидили-нозитолбисфосфат; ИФ3 – инозитол-1,4,5-трифосфат; ДАГ
– диацилглицерол; ЭР – эндоплазматический ретикулум; ФС – фосфодитилсерин. 1 –
взаимодействие адреналина с α1-рецептором трансформирует сигнал
через активацию G-белка на фосфолипазу С, переводя её в активное состояние; 2 –
фосфолипаза С гидроли-зует ФИФ2 на ИФ3 и ДАГ; 3 – ИФ3
активирует мобилизацию Са2+ из ЭР; 4 – Са2+, ДАГ и
фосфодитилсерин активируют протеинкиназу С. 5 – протеинкиназа С фосфорилирует
гликогенсинтазу, переводя её в неактивное состояние; 5 – комплекс 4Са2+-кальмодулин
активирует киназу фосфорилазы и кальмодулинзависимые протеинкиназы; 6 – киназа
фосфорилазы фосфорилирует гликогенфосфорилазу и тем самым её активирует; 7 –
активные формы трёх ферментов (кальмодулинзависимая протеинкиназа, киназа
фосфорилазы и протеин-киназа С) фосфорилируют гликогенсинтазу в различных
центрах, переводя её в неактивное состояние.
Какая система передачи сигнала в клетку
будет использована, зависит от типа рецепторов, с которыми взаимодействует
адреналин. Так, взаимодействие адреналина с β2-рецепторами
клеток печени приводит в действие аденилатциклазную систему. Взаимодействие же
адреналина с а1-рецепторами «включает» инозитолфосфатный механизм
трансмембранной передачи гормонального сигнала. Результат действия обеих систем
– фосфорилирование ключевых ферментов и переключение процессов с синтеза
гликогена на его распад. Следует отметить, что тип рецепторов, который в
наибольшей степени вовлекается в ответ клетки на адреналин, зависит от
концентрации его в крови.
В период
пищеварения преобладает влияние инсулина, так как инсулин-глюкагоновый индекс в
этом случае повышается. на обмен гликогена противоположно глюкагону. Инсулин снижает концентрацию
глюкозы в крови в период пищеварения, действуя на метаболизм печени следующим
образом:
• снижает
уровень цАМФ в клетках, фосфо-рилируя (опосредованно через Ras-путь) и тем
самым активируя фосфодиэстеразу цАМФ – фермент, гидролизующий цАМФ с
образованием АМФ. Механизм влияния инсулина на уровень цАМФ в клетке подробнее
будет изложен в разделе 11;
•
активирует (через Ras-путь) фосфопроте-инфосфатазу гранул гликогена,
которая дефосфорилирует гликогенсинтазу и таким образом её активирует. Кроме
того, фос-фопротеинфосфатаза дефосфорилирует и, следовательно, инактивирует
киназу фос-форилазы и гликогенфосфорилазу;
• индуцирует
синтез глюкокиназы, тем самым ускоряя фосфорилирование глюкозы в клетке.
Следует напомнить, что регуля-торным фактором в метаболизме гликогена является
также величина Кт глюкокиназы, которая много выше, чем Кт
гексокиназы. Смысл этих различий понятен: печень не должна потреблять глюкозу
для синтеза гликогена, если её количество в крови в пределах нормы.
Всё это вместе
приводит к тому, что инсулин одновременно активирует гликогенсинтазу и
ингибирует гликогенфосфорилазу, переключая процесс мобилизации гликогена на его
синтез.
В печени существует и аллостерическая
регуляция гликогенфосфорилазы, обеспечивающая внутриклеточные потребности в
глюкозе, но гормональные сигналы имеют приоритет над внутриклеточными и
преследуют другие физиологические цели. Ранее (см. раздел 6) рассматривалось
значение изменения в клетке уровней АТФ, АДФ и АМФ как показателя, отражающего
потребности клетки в энергии. Замедление утилизации АТФ сопровождается
снижением активности гликогенфосфорилазы и уменьшением скорости распада гликогена.
Напротив, увеличение расходования АТФ ведёт к повышению уровня АМФ, активации
гликогенфосфорилазы и ускорению распада гликогена. АТФ и АМФ являются
ал-лостерическими эффекторами по отношению к гликогенфосфорилазе. Существует
также и метаболический контроль активности гликогенфос-форилазы. Так, при
повышении концентрации глюкозо-6-фосфата активность этого фермента в клетках
печени снижается.
Имизм переваривания углеводов.
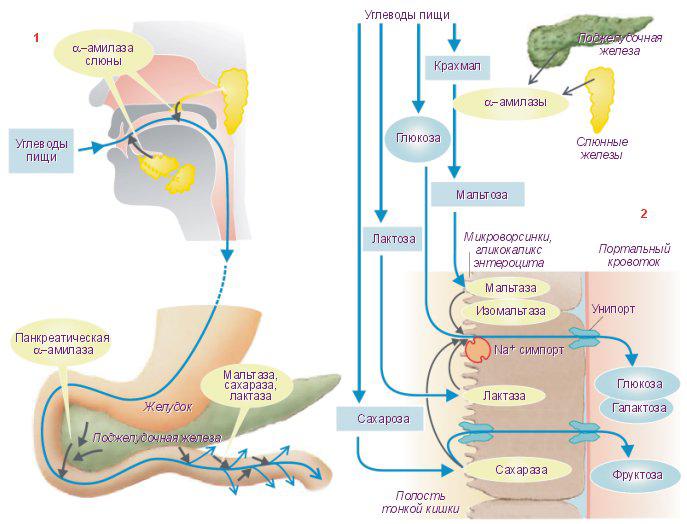
Ереваривание у разных видов животных.
У
моногастричных животных:
Первым этапом обмена является пищеварение,
в результате которого сложные питательные
вещества корма, превращаются в
низкомолекулярные, легкорастворимые
вещества, которые всасываются в кровь
и лимфу и используются организмом как
энергетический и строительный материал.
Пищеварение-
это процесс механических, биологических,
биохимических, биохимических изменений
корма в пищеварительном тракте. Углеводы,
которые поступают в организм животного-
растительный крахмал и представители
олиго и полисахаридами. По перевариваемости
подразделяют: -легкоперевариваемые
(крахмал, гликоген, энулин) –
трудноперевариваемые (целлюлоза,
клетчатка) – организм не вырабатывает
на них ферменты. В ротовой полости
углеводы начинают перерабатываться
(свинья, человек). Пища измельчается,
смачивается слюной (секрет слюнных
желез Слюна – жидкость без цвета, запаха,
тягучей консистенции, слегка мутная,
реакция слюны у всех животных разная –
слабощелочная. В слюне содержится белок
Муцин, значение слюны: фактор жевания
(смачивание), извлекает вкусовые в-ва,
поддерживает рецепторы, обволакивает
пищевой ком (способность скольжения
кома по пищеварительному тракту), Амилаза
– фермент, основаниями она нейтрализует
повышенную кислотность желудка. В
ротовой полости начинают расщепляться
крахмал, гликоген. Расщепление крахмала
идет ступенчато. В ротовой полости
крахмал полностью не переваривается,
потому что пища находится там
непродолжительное время, не у всех
животных есть амилаза в слюне. В желудке
моногастричных вырабатывается желудочный
сок, рН – резкокислый, 1,5 – 2,5. Ферменты
в такой среде не вырабатываются.
Окончательное переваривание происходит
в 12ти перстной кишке, в неё открывается
проток поджелудочной железы, оны
вырабатывает секрет, который содержит
ферменты на белки, жиры и углеводы. На
углеводы – амилаза, декстриназа,
мальтаза, сахараза и т.д. 1 и 2 расщепляет
крахмал, путем реакции гидролиза до
мальтозы – мальтазы – 2-е молекулы
альфа, дэ глюкозы. Под действием ферментов
углеводы расщепляются, всасываются в
тонком кишечнике в виде фосфорных
эфиров. Конечный продукт расщепления
углеводов у однокамерных- глюкоза,
образуется в тонком кишечнике.
Троение и роль в организме гепарина, гиалуроновой, хондроитинсерной кислоты.
Гепарин
– природный полисахарид. Содержится в
тканях млекопитающих: печени, легких,
селезенке, почках, коже, в стенках
кровеносных сосудов; синовиальной
жидкости суставов; стекловидном теле
глаза. В живом организме гепарин в
свободном состоянии не встречается,
поскольку всегда связан с молекулами
белка, образуя так называемый
углевод-белковый комплекс (протеогликан).
Совместно с фибринолизином он входит
в состав физиологической антисвертывающей
системы. Введение гепарина в организм
сопровождается понижением содержания
холестерина в сыворотке крови, улучшением
коронарного кровотока.
Гиалуроновая
кислота
– природная смазка для суставов. В самом
простом понимании – сахар. Благодаря
своей структуре и большой молекулярной
массе ее молекулы способны удерживать
количество огромное количество воды.
Хондроитинсерная
кислота
– относится к протеогликанам. Протеогликаны,
содержащие хондроитинсерные кислоты,
входят в состав межклеточного вещества
соединительной ткани позвоночных
животных, в том числе человека, кроме
того, протеогликаны обнаружены в коже,
костной и хрящевой тканях, стекловидном
теле и роговице глаза, сосудистой стенке,
тканях сердца и мозга. Биологическое
значение хондроитинсерных кислот
состоит в их влиянии на водно-солевой
обмен благодаря способности связывать
воду и катионы, регулировать процесс
диффузии. Вследствие того, что
хондроитинсерные кислоты являются
вязкими и в значительной степени
гидратированными полианионами, они
препятствуют сжатию соединительной
ткани и играют роль своеобразной смазки
суставных поверхностей.






